Präsentation herunterladen
Die Präsentation wird geladen. Bitte warten

1
Entwicklungsneurobiologie
IV. Musterbildung WS 2008/09

2
Differenzierungsspotential der Vorläuferzellen wird
eingeschränkt während der Entwicklung Regionale Unterschiede Regionale Identität

3
Neurale Induktion als Hemmung der Differenzierung des Ektoderms zu Epidermis

4
Zellen aus dem vorderen Teil der induzierten Neuralplatte (schwarz) werden mit Zellen
aus dem posterioren Notochord (gelb) gemischt in verschiedenen Verhältnissen: Posteriore Markergene (Hindbrain, Spinal cord) werden durch Notochord induziert.
 gemischt in verschiedenen Verhältnissen: Posteriore Markergene (Hindbrain, Spinal cord) werden durch Notochord induziert.")
5
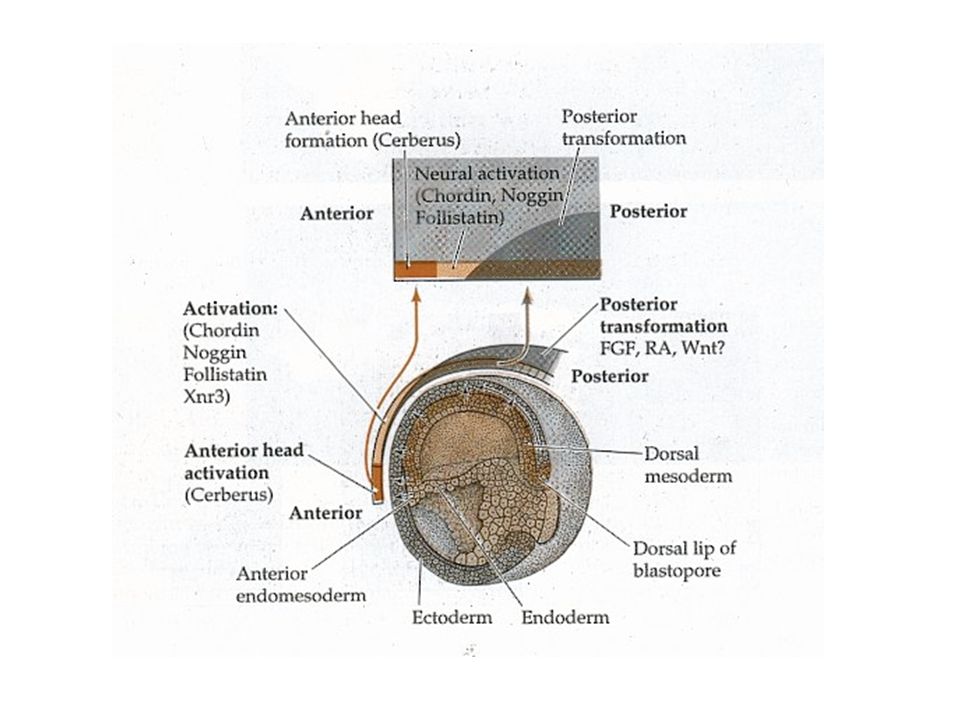
Aktivation-Transformation des Neuroepithels

7
Anterior-posteriore (A/P) Musterbildung
 Musterbildung")
8
Musterbildung und Morphogen-Gradienten
Morphogene spezifizieren direkt unterschiedliche Zell- Schicksale durch unterschiedliche Morphogen-Konzentrationen

9
Musterbildung und Morphogen-Gradienten
Gradientenbildung und Aufrechterhaltung: Bestimmt durch Produktion, Diffusion und Degradation Diffusion erlaubt Gradienten über Felder von 50 Zellen (F. Crick) Effektive Difusionsraten sind sehr klein Degradation wird z.T. durch das Morphogen stimuliert (feed-f.)
 Effektive Difusionsraten sind sehr klein. Degradation wird z.T. durch das Morphogen stimuliert (feed-f.)")
10
Musterbildung und Morphogen-Gradienten
Interpretation von Gradienten: (L. Wolpert: French Flag)
")
11
Musterbildung und Morphogen-Gradienten
Interpretation von Gradienten: Es können zwischen 3-7 Schwellenwerte (Zellschicksale) durch einen Gradienten bestimmt werden Es werden kleine, 2-3-fache Konzentrationsunterschiede detektiert Die Signaltransduktion ist linear (derselbe Rezeptor, derselbe Signalweg, derselbe Transkriptionsfaktor bei unterschiedlichen Konzentrationen) Morphogenkonzentration wird in Transkriptionsfaktorkonzentration über- setzt (BMP4 in Smad2; Shh in Gli)
 durch einen. Gradienten bestimmt werden. Es werden kleine, 2-3-fache Konzentrationsunterschiede detektiert. Die Signaltransduktion ist linear (derselbe Rezeptor, derselbe Signalweg, derselbe Transkriptionsfaktor bei unterschiedlichen Konzentrationen) Morphogenkonzentration wird in Transkriptionsfaktorkonzentration über- setzt (BMP4 in Smad2; Shh in Gli)")
12
Musterbildung und Morphogen-Gradienten
Interpretation von Gradienten: 5. Unterschiedliche Gen-Promotoren besitzen unterschiedliche Affinitäten für einen bestimmten Transcriptionsfaktor

13
Musterbildung und Morphogen-Gradienten
Interpretation von Gradienten: 6. Unterschiedliche Gen-Promotoren besitzen unterschiedliche Affinitäten für einen bestimmten Repressor

14
Musterbildung und Morphogen-Gradienten

15
Musterbildung und Morphogen-Gradienten
Fehlerkorrektur:

16
1. Beispiel für A/P Musterbildung
Wnt kontrolliert A/P Musterbildung im Vorder-Hinterhirnbereich

17
Wnt kontrolliert A/P Musterbildung im Vorder-Hinterhirnbereich
(zusammen mit FGF und RA) - kaudalisierender Effekt Frz bead in a: unter zukünft. MB Wnt bead in b: unter zukünft. FB
 - kaudalisierender Effekt. Frz bead in a: unter zukünft. MB. Wnt bead in b: unter zukünft. FB.")
18
Anterior-posteriore (A/P) Musterbildung im Hinterhirn
 Musterbildung im Hinterhirn")
19
Rhombomeren sind Kompartimente

20
Rhombomeren sind verschieden bzgl. Funktion ihrer Neuronen
trochlear n.trigeminal n.facialis n.glossopharyngeus n.vagus n.hypoglossus

21
Wie wird die A/P-Identität der Rhombomeren bestimmt?

22
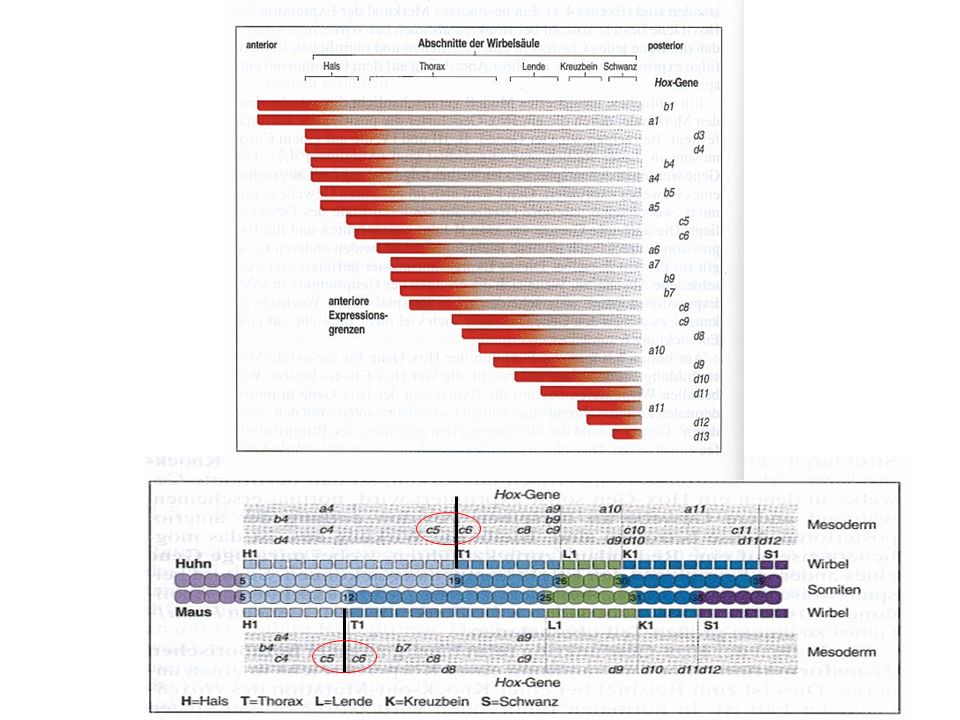
Homöobox-Genfamilie (Hox-Komplexe)
")
24
Welchen Effekt hat die Eliminierung von z.B. Hoxb1?
Transplantation von r4 nach r2 bei stage 9 A/P Identität und Hox code gleichzeitig früh fest gelegt (stage 6-8), bevor sich Neuralrohr schließt Welchen Effekt hat die Eliminierung von z.B. Hoxb1?
, bevor sich. Neuralrohr schließt. Welchen Effekt hat die Eliminierung von z.B. Hoxb1")
25
Hoxb1 ist essentiell für die Entstehung von r4-Identität
Wie wird die Expression von Hox-Genen im Hinterhirn kontrolliert?

26
Kontrolle der Expression von Hox-Genen im Hinterhirn
durch Retinsäure Wenn Retinsäure entfernt wird, fehlen r4-7, r1-3 vergrößert Retinsäure wird lokal in cervikalem paraxialem Mesoderm (Somiten) Produziert, diffundiert ins Neuralrohr und steuert Hox-Genexpression Retinsäure führt zur A/P Transformation - Rhombencephalon vergrößert Mittelhirn und Vorderhirn kleiner. Retinsäure ist ein Derivat von Vitamin A; Aktiviert sog. Kern- Rezeptoren; ist bei vielen Musterbildungsprozessen beteiligt z.B. A/P Polarität der Extremitäten
 Produziert, diffundiert ins Neuralrohr und. steuert Hox-Genexpression. Retinsäure führt zur A/P Transformation - Rhombencephalon vergrößert. Mittelhirn und Vorderhirn kleiner. Retinsäure ist ein Derivat von Vitamin A; Aktiviert sog. Kern- Rezeptoren; ist bei vielen Musterbildungsprozessen beteiligt. z.B. A/P Polarität der Extremitäten.")
27
Anterior-posteriore (A/P) Musterbildung im Rückenmark
Unterschiedliche Typen von Motoneuronen im Rückenmark: Funktion und Lokalisation P MMC CT A LMC A/P Identität von Motoneuronen im RM durch Hox-Gene bestimmt

28
Hox-Expression wird im Neuralplattenstadium festgelegt

29
Induktion der Hox gene Expression durch verschiedene FGF-Konzentrationen
5 ng/ml FGF brachial 25 ng/ml FGF thorakal 125 ng/ml FGF lumbar 625 ng/ml FGF P

30
Jeder Muskel (50) wird von einer spezifischen Motoneurongruppe (pool) innerviert
A brachial

31
Hox5/Hoxc8 bestimmen die Identität von Motoneuronpools (A/P) in LMCbrachial
 in LMCbrachial")
32
Dorsoventrale Musterbildung

33
Dorsoventrale Musterbildung im Rückenmark

34
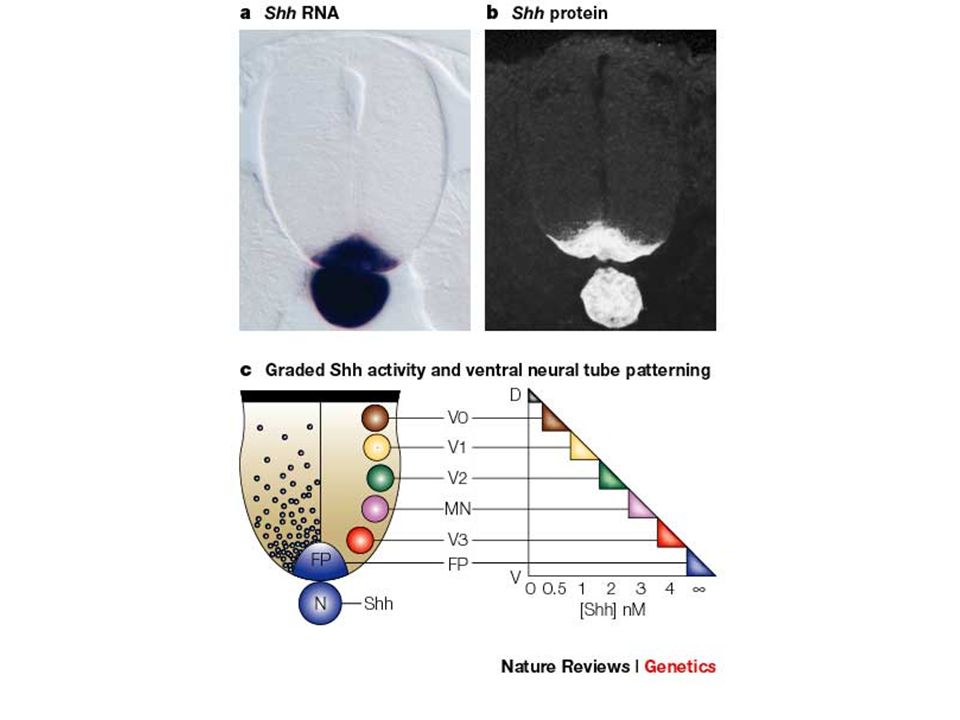
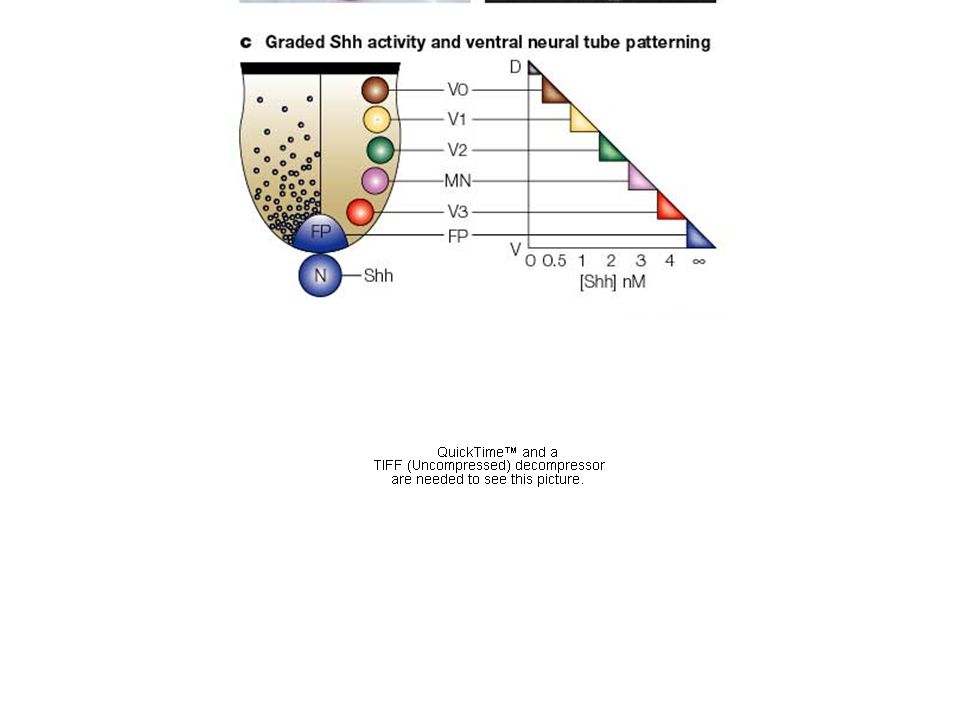
Das ventralisierende Signal aus dem Notochord ist ein
sekretiertes Protein: sonic hedgehog (shh)
")
35
Sonic hedgehog (shh) Signaltransduktion
 Signaltransduktion")
37
Shh reprimiert Pax3,7 > Pax6
Typ I Homöobox-Gene und induziert Typ II ...

38
Definition von Bereichen mit unterschiedlicher Homöodomänen-Genexpression
In den unterschiedlichen Bereichen entstehen verschiedene ventrale Neuronen

39
Nicht nur die Konzentration sondern auch die Dauer ist wichtig
Shh ist ein Morphogen „Identische Vorläuferzellen differenzieren in Abhängigkeit von der Konzentration eines Morphogens zu unterschiedlichen Zelltypen“ Nicht nur die Konzentration sondern auch die Dauer ist wichtig (Dessaud et al., (2007) Nature 450, )
 Nature 450, )")
41
Dorsalisierende Signale im ZNS

42
Im dorsalen Rückenmark entstehen Neuralleiste, Deckplatte (roof plate) und dorsale Interneuronen (C)
Die Entstehung erfordert dorsalisierende induktive Signale. Bone morphogenetic proteins (BMPs) werden im epidermalen Ektoderm exprimiert und induzieren Neuralleiste (slug). Dorsale Interneuronen enstehen später unter dem Einfluß von Signalen (BMPs) aus der Deckplatte.
 werden im epidermalen Ektoderm exprimiert und induzieren Neuralleiste (slug). Dorsale Interneuronen enstehen später unter dem Einfluß von Signalen (BMPs) aus der Deckplatte.")
43
Musterbildung im dorsalen Rückenmark
Unterschiedliche neuronale Phänotypen werden durch unterschiedliche Konzentrationen von BMPs (A) als auch selektiv durch verschiedene Mitglieder der BMP Familie induziert (B)
 als auch selektiv. durch verschiedene Mitglieder der BMP Familie induziert (B)")
44
GDF-7 wird in der Deckplatte exprimiert.
Im GDF-7 knockout verschwindet die mATH1- Expression und D1A Interneurone entstehen nicht. D1B Neurone sind nicht betroffen.

45
Eliminierung der Deckplatte durch Diphterietoxin-GDF7: Beweis dass die Entwicklung der dorsalen Interneurone D1A, D1B und D2 von Signalen aus der Deckplatte bestimmt wird.

47
Integration der A/P- und D/V Positionsinformation
ektopisch shh shh ektopische dopaminerge Neuronen dopaminerge Neuronen serotoninerge Neuronen Motoneuronen

48
Musterbildung im Mittelhirn-Vorderhirn

49
Prosomeren im Mittel- und Vorderhirnbereich der Neuralplatte zu erkennen durch die Expression von Transkriptionsfaktoren

50
A/P und D/V Signale auch im Mittel-, Zwischen- und Telencephalon
BMP in der MHB FGF Shh BMP Shh

51
Zusammenfassung I A/P Musterbildung durch Gradienten sekretierter Faktoren mit kaudalisierender Funktion (Vorderhirn Wnt, Rautenhirn Retinsäure, Rückenmark FGF) A/P Muster durch Hox-Gene im Rückenmark und Rautenhirn (durch FGF und Retinsäure induziert) D/V Musterbildung durch Gradienten sekretierter Faktoren (shh, BMPs) mit ventralisierender/dorsalisierender Funktion Identische D/V Signale im Rückenmark, Hinterhirn, Mittelhirn und Endhirn (shh, BMPs).
 A/P Muster durch Hox-Gene im Rückenmark und Rautenhirn. (durch FGF und Retinsäure induziert) D/V Musterbildung durch Gradienten sekretierter Faktoren. (shh, BMPs) mit ventralisierender/dorsalisierender Funktion. Identische D/V Signale im Rückenmark, Hinterhirn, Mittelhirn und Endhirn (shh, BMPs).")
52
Zusammenfassung II D/V Muster durch Homöoprotein-Domänen (Pax6, Dlx ...) durch shh, BMPs induziert. Dorsale Signale aus Ektoderm, ventrale aus Mesoderm, später beide ins Neuroektoderm verlagert (Boden- und Deckplatte). D/V Signale kommen nach A/P Muster 3D-Koordinatensystem. das die regionale Identität definiert.
. D/V Signale kommen nach A/P Muster. 3D-Koordinatensystem. das die regionale Identität definiert.")
54
MHB ist ein ‚Signalzentrum‘ wie Bodenplatte/Deckplatte
(FGF8 Applikation zeigt denselben Effekt)
")
56
BMPs sind für die Entstehung dorsaler Phänotypen im ZNS verantwortlich
A) Frühes Stadium: B) Späteres Stadium:
 Frühes Stadium: B) Späteres Stadium:")
Ähnliche Präsentationen













>")








