Präsentation herunterladen
Die Präsentation wird geladen. Bitte warten

1
Studienordnung für das Fach Neurbiologie:
HF NF + + Vorlesungen 6 SWS + + F-I Praktikum 12 SWS + F-II Praktikum 20 SWS + Spezialpraktikum 8 SWS + + Genetische Übungen 2 SWS + + Genetisches Seminar 2 SWS F-I Praktikum: 3 Blöcke Block I: Wahl zw. Neurogenetik und Tierphysiologie Block II: Wahl zw. Klin. Neurobiologie und Neurologie Block III: Wahl zw. Anatomie, Physiologie und Psychiatrie Wissensnachweis für Aufnahme ins F-I Praktikum : Klausur über Stoff der Vorlesungen Mol. Neurobiologie und Klinische Neurobiologie am Fr. 1. Februar Uhr Nachklausur (falls genügend Plätze vorhanden) am 22. Februar Uhr HS A102. Praktikumstermine (ab Januar): werden durch Aushang bekannt gegeben Humangenetik 2x 2 SWS, Immungenetik, Virusgenetik, Entwicklungsgenetik
 am 22. Februar Uhr HS A102. Praktikumstermine (ab Januar): werden durch Aushang bekannt gegeben. Humangenetik 2x 2 SWS, Immungenetik, Virusgenetik, Entwicklungsgenetik.")
2
Kap. I ELEKTRISCHE GRÖSSEN
Ladung Q, Einheit Coulomb [CB], Elementarladung = Ladung des Elektrons e = 1,6 x [Cb] Alle Ladungen sind Vielfaches von e. Zwischen Ladungen treten Kräfte auf: Gleiche Ladungen stoßen sich ab, ungleiche ziehen sich an. Jede Ladung erzeugt ein elektrisches Feld. Feldstärke E, Einheit Volt/Meter [V/m] oder Newton/Coulomb [N/Cb]. Das Feld übt auf eine (kleine) Testladung q eine Kraft aus: K = q x E Das Feld lässt sich durch durch Feldlinien beschreiben, die Kraft wirkt entlang der Feldlinien, die Dichte der Feldlinien ist ein Maß für die Feldstärke. Die Feld- linien verlaufen von positiven zu negativen Ladungen. Bewegt man eine (positive) Testladung gegen den Verlauf der Feldlinien, so verrichtet man Arbeit und erhöht die potentielle Energie der Testladung. Die potentielle Energie pro Ladungseinheit nennt man das Potential U, Einheit Volt [V]. [V] = [N x m]/[Cb] Konvention: Im Unendlichen ist das Potential U = 0. Beispiel: Punktladung Q bei r = 0: U Spannung U(r) ~ Q/r r
 Testladung q eine Kraft aus: K = q x E. Das Feld lässt sich durch durch Feldlinien beschreiben, die Kraft wirkt entlang der Feldlinien, die Dichte der Feldlinien ist ein Maß für die Feldstärke. Die Feld- linien verlaufen von positiven zu negativen Ladungen. Bewegt man eine (positive) Testladung gegen den Verlauf der Feldlinien, so verrichtet man Arbeit und erhöht die potentielle Energie der Testladung. Die potentielle Energie pro Ladungseinheit nennt man das. Potential U, Einheit Volt [V]. [V] = [N x m]/[Cb] Konvention: Im Unendlichen ist das Potential U = 0. Beispiel: Punktladung Q bei r = 0: U. Spannung. U(r) ~ Q/r. r.")
3
Eine Potentialdifferenz bezeichnet man auch als Spannung.
Entlang der Feldlinien ändert sich das Potential am stärksten, senkrecht zu ihnen ändert es sich nicht. Ändert sich das Potential z.B. nur in x - Richtung, so ist E = - Eine Potentialdifferenz bezeichnet man auch als Spannung. Eine Spannung an den Enden eines schlechten Leiters (Widerstands) führt zum Strom I, Einheit Ampère [A] Der Strom ist gleich der pro Zeiteinheit verschobenen Ladungen: I = [A] = [Cb]/[s] Strom und Spannung am Widerstand R, Einheit Ohm [W], folgen dem Ohm‘schen Gesetz: I = U/R U = R x I R = U/I [W] = [V]/[A] Leitfähigkeit G, Einheit Siemens [S] ist inverser Widerstand G = 1/R [S] = [mho] = [W-1] dU dx dQ dt Beispiel
 führt zum. Strom I, Einheit Ampère [A] Der Strom ist gleich der pro Zeiteinheit verschobenen Ladungen: I = [A] = [Cb]/[s] Strom und Spannung am. Widerstand R, Einheit Ohm [W], folgen dem Ohm‘schen Gesetz: I = U/R U = R x I R = U/I [W] = [V]/[A] Leitfähigkeit G, Einheit Siemens [S] ist inverser Widerstand. G = 1/R [S] = [mho] = [W-1] dU. dx. dQ. dt. Beispiel.")
4
Fläche f Länge l R = r x l/f r = spezifischer Widerstand [W x m]
Beispiel Draht: Fläche f Länge l R = r x l/f r = spezifischer Widerstand [W x m] Spezifische Widerstände: z.B. Zylinder Länge 1 cm, Durchmesser 1 mm Kupfer: r = 1,7 x 10-8 [W x m] R = 0,2 mW (Milliohm) Axoplasma: r = 0,3 [W x m] R = 3,8 kW Membranmaterial: r = 108 [W x m] R = 1,27 x 1012 W Glas: r = 5 x 1016 [W x m] R = 6,4 x 1020 W Für eine Membran spielt neben dem Widerstand die Kapazität C, Einheit Farad [F] [F] = [Cb/V] eine wichtige Rolle. Bringt man eine Ladung Q auf eine leitende Kugel, so ist das Potential U der Kugeloberfläche (gegenüber unendlich) proportional zur Ladungsmenge: U = Q/C Q = C x U Plattenkondensator, 2 Platten der Fläche F mit Abstand a: C proportional zu F/a Da gleichartige Ladungen sich gegenseitig abstoßen, wird das Potential vom Radius der Kugel abhängen. Bei gleicher Ladung wird eine kleine Kugel ein höheres Potential haben als eine große, d.h. eine kleine Kugel hat eine kleine Kapazität, eine große eine große Kapazität. Bei einem Plattenkondensator ist die Kapazität proportional zur Fläche und umgekehrt proportional zum Abstand a der Platten.
![Fläche f Länge l R = r x l/f r = spezifischer Widerstand [W x m]](http://slideplayer.org/slide/205817/1/images/4/Fl%C3%A4che+f+L%C3%A4nge+l+R+%3D+r+x+l%2Ff+r+%3D+spezifischer+Widerstand+%5BW+x+m%5D.jpg "Beispiel Draht: Fläche f Länge l R = r x l/f r = spezifischer Widerstand [W x m] Spezifische Widerstände: z.B. Zylinder Länge 1 cm, Durchmesser 1 mm. Kupfer: r = 1,7 x 10-8 [W x m] R = 0,2 mW (Milliohm) Axoplasma: r = 0,3 [W x m] R = 3,8 kW. Membranmaterial: r = 108 [W x m] R = 1,27 x 1012 W. Glas: r = 5 x 1016 [W x m] R = 6,4 x 1020 W. Für eine Membran spielt neben dem Widerstand die. Kapazität C, Einheit Farad [F] [F] = [Cb/V] eine wichtige Rolle. Bringt man eine Ladung Q auf eine leitende Kugel, so ist das Potential U der. Kugeloberfläche (gegenüber unendlich) proportional zur Ladungsmenge: U = Q/C Q = C x U. Plattenkondensator, 2 Platten der Fläche F mit Abstand a: C proportional zu F/a. Da gleichartige Ladungen sich gegenseitig abstoßen, wird das Potential. vom Radius der Kugel abhängen. Bei gleicher Ladung wird eine kleine. Kugel ein höheres Potential haben als eine große, d.h. eine kleine Kugel. hat eine kleine Kapazität, eine große eine große Kapazität. Bei einem. Plattenkondensator ist die Kapazität proportional zur Fläche und. umgekehrt proportional zum Abstand a der Platten.")
5
Die Axon-Membran zwischen Extrazellulärflüssigkeit und Axoplasma wirkt als
Plattenkondensator. Da die Membrandicke relativ konstant ist, gibt man die Kapazität pro Fläche als spezifische Membrankapazität cm an. Es gilt: C = cm x f Für biologische Membranen ist cm = 1 µF/cm2 Da Strom gleich Ladungsverschiebung pro Zeiteinheit ist, IC = dQ/dt = C x dU/dt fließt durch einen Kondensator nur ein Strom, wenn sich die Spannung mit der Zeit ändert, d.h. dU/dt = 0 ist.

6
Kap. II Einfache lineare Netzwerke:
1) Zwei Widerstände R1 und R2 hintereinander („in Serie“) geschaltet haben den Gesamtwiderstand Rges = R1 + R2 Zwei Widerstände parallel geschaltet: Rges = R1 x R2/(R1 + R2) (Bei Kondensatoren umgekehrt) 2) Ein Axonabschnitt verhält sich in erster Näherung wie ein Tiefpass: R.C = t = „Zeitkonstante“ des Tiefpasses E(t) = Eingangssignal (bekannt) UC(t) = gesuchtes Ausgangssignal R R2 R UR + UC = E UR = I x R I = C x dUC/dt R.C x dUC/dt + UC = E E(t) UC
 Zwei Widerstände R1 und R2 hintereinander („in Serie ) geschaltet haben den Gesamtwiderstand Rges = R1 + R2. Zwei Widerstände parallel geschaltet: Rges = R1 x R2/(R1 + R2) (Bei Kondensatoren umgekehrt) 2) Ein Axonabschnitt verhält sich in erster Näherung wie ein Tiefpass: R.C = t = „Zeitkonstante des Tiefpasses. E(t) = Eingangssignal (bekannt) UC(t) = gesuchtes Ausgangssignal. R1 R2. R. UR + UC = E UR = I x R I = C x dUC/dt. R.C x dUC/dt + UC = E. E(t) UC.")
7
Beispiel: E(t) = Stufenfunktion = 0 für t < 0 = U0 für t > 0
Diese „lineare Differentialgleichung erster Ordnung“ ist für viele Funktionen E(t) leicht lösbar. Beispiel: E(t) = Stufenfunktion = 0 für t < 0 = U0 für t > 0 U(t) = U0.(1 – e-t/t) In linearen Netzwerken gilt das „Superpositionsprinzip“: Ist U1(t) die Antwort des Netzwerks bei Eingangssignal E1(t) und U2(t) Antwort bei Eingangssignal E2(t), so ist U1(t) + U2(t) die Antwort bei Eingangssignal E1(t) + E2(t). Lineare Netzwerke bestehen nur aus linearen Komponenten wie z.B. Widerständen, Kondensatoren, Verstärkern, Integratoren, Differentiatoren, Addierern, Subtrahierern. Nichtlineare Komponenten sind z.B. Gleichrichter, Multiplikatoren, adaptive Komponenten. E(t) U0 U(t) t Beispiel
 leicht. lösbar. Beispiel: E(t) = Stufenfunktion. = 0 für t < 0. = U0 für t > 0. U(t) = U0.(1 – e-t/t) In linearen Netzwerken gilt das „Superpositionsprinzip : Ist U1(t) die Antwort des Netzwerks bei Eingangssignal E1(t) und U2(t) Antwort bei. Eingangssignal E2(t), so ist U1(t) + U2(t) die Antwort bei Eingangssignal E1(t) + E2(t). Lineare Netzwerke bestehen nur aus linearen Komponenten wie z.B. Widerständen, Kondensatoren, Verstärkern, Integratoren, Differentiatoren, Addierern, Subtrahierern. Nichtlineare Komponenten sind z.B. Gleichrichter, Multiplikatoren, adaptive. Komponenten. E(t) U0. U(t) t. Beispiel.")
8
= + = E(t) U0 2. Beispiel: Rechteckimpuls t E(t) E(t) U0 U0 U0 U(t) t
Nadelimpuls = t E(t) Die Antwort kann mit Hilfe des Superpositionsprinzips ermittelt werden.
 Die Antwort kann mit Hilfe des Superpositionsprinzips ermittelt werden.")
9
3. Beispiel: Nadelimpuls (Rechteck mit U0 , Dt 0), U0 x Dt = A o o
Impulsantwort des Tiefpasses: U(t) = (A/t) x e-t/t o o E(t) A/t Sinusfunktion t t E(t) U(t)
 = (A/t) x e-t/t. o. o. E(t) A/t. Sinusfunktion. t. t. E(t) U(t)")
10
4. Beispiel: E(t) = sin (wt) Antwort: U(t) = A sin(wt + F(w))
(w = „Kreisfrequenz“ = 2pn n = Frequenz = Anzahl der Schwingungen/sec) Die Antwort des Tiefpasses auf ein sinusförmiges Eingangssignal ist wieder eine Sinusfunktion mit gleicher Frequenz, aber veränderter Amplitude A und verschobener Phase F. A und F sind Funktionen von w. Dies gilt für alle linearen Netzwerke!
 Die Antwort des Tiefpasses auf ein sinusförmiges Eingangssignal ist. wieder eine Sinusfunktion mit gleicher Frequenz, aber veränderter. Amplitude A und verschobener Phase F. A und F sind Funktionen von w. Dies gilt für alle linearen Netzwerke!")
11
A(w) = Amplitudenfrequenzgang, doppelt-logarithmische Darstellung
F(w) = Phasenfrequenzgang, halb-logarithmische Darstellung (Bode-Diagramm) w o Knick bei w = 2pn = 1/t o w o
 = Phasenfrequenzgang, halb-logarithmische Darstellung (Bode-Diagramm) w. o. Knick bei w = 2pn = 1/t. o. w. o.")
12
Da man jede beliebige Eingangsfunktion als Summe
von Sinusfunktionen darstellen kann (Fourier-Reihe bzw. –Integral), kann man in vielen Fällen das Ausgangs- signal mit Hilfe des Superpositionsprinzips berechnen. Für die beiden wichtigsten linearen Filter (Hochpass und Tiefpass) sind die Eingangs-Ausgangs-Beziehungen für Spannungsstufe, Rechteckimpuls, Nadelimpuls und Sinus-Funktion noch mal gegenübergestellt.
, kann man in vielen Fällen das Ausgangs- signal mit Hilfe des Superpositionsprinzips berechnen. Für die beiden wichtigsten linearen Filter (Hochpass. und Tiefpass) sind die Eingangs-Ausgangs-Beziehungen. für Spannungsstufe, Rechteckimpuls, Nadelimpuls. und Sinus-Funktion noch mal gegenübergestellt.")
13
Hochpass Tiefpass R x C = t Eingangsfunktion Ausgangsfunktionen

14
Hochpass Tiefpass

15
Hochpass Tiefpass

16
Kap. III Ionale Grundlagen des Ruhepotentials
Bernstein-Theorie (1902): Annahme: Membran nur für K+ durchlässig, dann gilt die Nernst‘sche Gleichung: U0 = (RT/F) x ln([K+]a/[K+]i Chemische Analysen: K+ innen hoch, Na+ innen niedrig, gleiche physikalische Grundlagen für beide: Ionenkonzentrationsgradient über semipermeabler Membran. Genauer betrachten. Skizze. K = mb + rv Erst 40 Jahre später: Goldman-Hodgekin-Katz Gleichung (Membran für Na+, K+ und Cl- durchlässig)
: Annahme: Membran nur für K+ durchlässig, dann gilt die Nernst‘sche Gleichung: U0 = (RT/F) x ln([K+]a/[K+]i. Chemische Analysen: K+ innen hoch, Na+ innen niedrig, gleiche physikalische Grundlagen für beide: Ionenkonzentrationsgradient über semipermeabler Membran. Genauer betrachten. Skizze. K = mb + rv. Erst 40 Jahre später: Goldman-Hodgekin-Katz Gleichung. (Membran für Na+, K+ und Cl- durchlässig)")
17
Bewegungsgleichung für geladene Teilchen in wässriger Lösung
K = m x b + (1/w) x v = m x (dv/dt) + (1/w) x v m = Masse des Teilchens, b = Beschleunigung, w = Beweglichkeit des Teilchens im Lösungsmittel, (1/w = Reibungsfaktor), v = Geschwindigkeit 2 Arten von Kräften, makroskopische Prozesse Diffusion, Elektrophorese. Reibung in Lösungsmittel: Bewegungsgleichung. je leichter, desto schneller Endgeschwindigkeit. Für kleine Ionen in wässriger Lösung Beschleunigungsphase < µsec, vernachlässigbar. K = (1/w) x v t = m/w < 1 psec Näherung:
 x v = m x (dv/dt) + (1/w) x v. m = Masse des Teilchens, b = Beschleunigung, w = Beweglichkeit des Teilchens im Lösungsmittel, (1/w = Reibungsfaktor), v = Geschwindigkeit. 2 Arten von Kräften, makroskopische Prozesse Diffusion, Elektrophorese. Reibung in Lösungsmittel: Bewegungsgleichung. je leichter, desto schneller Endgeschwindigkeit. Für kleine Ionen in wässriger Lösung Beschleunigungsphase < µsec, vernachlässigbar. K = (1/w) x v. t = m/w < 1 psec Näherung:")
18
2 Kräfte: Diffusion und Elektrophorese
Diffusion: kleine Teilchen in wässriger Lösung: wie Gas. pV = nRT n = Anz. Mole, R = allg. Gaskonstante, T = abs. Temp (°K) 1. Fick‘sches Gesetz: JD(x) = - D x dc(x)/dx D = wRT/L = Diff.-Konst. J = Ionenfluss c = Ionenkonzentration L = Loschmidtsche Zahl Elektrophorese: JE(x) = - (zDF/RT) x dU(x)/dx z = Wertigkeit des Ions U = Spannung F = Faraday Konstante J = JD + JE = -D(dc/dx + (zF/RT) x c x dU/dx) Bernstein-Theorie: Membran nur für K+ durchlässig Gleichgewichtszustand: iK = 0 Nernst-Gleichung U0 = (RT/F) x ln([K+]a/[K+]i
 1. Fick‘sches Gesetz: JD(x) = - D x dc(x)/dx D = wRT/L = Diff.-Konst. J = Ionenfluss c = Ionenkonzentration L = Loschmidtsche Zahl. Elektrophorese: JE(x) = - (zDF/RT) x dU(x)/dx z = Wertigkeit des Ions. U = Spannung F = Faraday Konstante. J = JD + JE = -D(dc/dx + (zF/RT) x c x dU/dx) Bernstein-Theorie: Membran nur für K+ durchlässig. Gleichgewichtszustand: iK = 0. Nernst-Gleichung U0 = (RT/F) x ln([K+]a/[K+]i.")
19
Tintenfisch Riesenaxon: (Konzentrationen in mM)
Axoplasma Meerwasser K Na Cl org. Anionen Ui = - 93 mV (bei 20°C) Axon 1 mm lang, 100µ Durchmesser: 109 K+-Ionen müssen rausdiffundieren, um das Ruhepotential aufzubauen von insgesamt 1015 d.h. nur 0,0001%
 Axon 1 mm lang, 100µ Durchmesser: 109 K+-Ionen müssen rausdiffundieren, um das Ruhepotential aufzubauen. von insgesamt 1015 d.h. nur 0,0001%")
20
Was leistet die Bernstein‘sche Theorie?
Aufrechterhaltung des Ruhepotentials ohne Energiezufuhr Vorzeichen und ungefähre Größe korrekt Erklärt die Erniedrigung des Ruhepotentials bei Erhöhung von [K+]a Wo versagt die Bernstein‘sche Theorie? Erklärt nicht Überschwingen des Nervenimpulses Bei niedrigen externen Kaliumkonzentrationen Abweichung von Theorie Misst man Natrium und Kalium Transportraten (mit Radioisotopen), so findet man etwa gleich große Werte Realistisch: iNa und iCl nicht 0. Gleichgewichtszustand: iges = iK + iNa + ICl = 0 Goldman-Hodgkin-Katz-Gleichung („Konstantfeldtheorie“) Um = (RT/F) x ln Tintenfisch-Riesenaxon: PK : PNa : PCl = 1 : 0,04 : 0,05 P-Na groß Um E-Na etc. Gleichgewichtspotential PNa [Na+]a + PK [K+]a + PCl [Cl-]i PS = DSbS/d PS = Permeabilität für Ionenart S b = Verteilungskoeff. d = Membrandicke PNa [Na+]i + PK [K+]i + PCl [Cl-]a
, so findet man. etwa gleich große Werte. Realistisch: iNa und iCl nicht 0. Gleichgewichtszustand: iges = iK + iNa + ICl = 0. Goldman-Hodgkin-Katz-Gleichung („Konstantfeldtheorie ) Um = (RT/F) x ln. Tintenfisch-Riesenaxon: PK : PNa : PCl = 1 : 0,04 : 0,05 P-Na groß Um E-Na etc. Gleichgewichtspotential. PNa [Na+]a + PK [K+]a + PCl [Cl-]i. PS = DSbS/d. PS = Permeabilität. für Ionenart S. b = Verteilungskoeff. d = Membrandicke. PNa [Na+]i + PK [K+]i + PCl [Cl-]a.")
21
„shunting inhibition“
Depolarisation: PNa vergrößern PK verkleinern Hyperpolarisation: PNa verkleinern PK vergrößern Wirkung von PCl: „shunting inhibition“ Beispiele: a) exzitat. Synapse, ACh auf Skelettmuskel, Photorezeptoren bei Invertebraten c) Photorezeptoren von Vertebraten d) ACh auf Herzmuskel, Photorezeptoren von Pecten
 exzitat. Synapse, ACh auf Skelettmuskel, Photorezeptoren bei Invertebraten. c) Photorezeptoren von Vertebraten. d) ACh auf Herzmuskel, Photorezeptoren von Pecten.")
22
Strom-Spannungs Beziehungen am passiven Axon (Konstant-Feld Theorie)
Der Strom für die Ionenart „S“ ist proportional zur Permeabilität PS der Membran für diese Ionenart, hängt aber in komplexer Weise ab von Um, den Ionenkonzentrationen [S] innen und außen und der abs. Temperatur T Übergang zu Bernstein-Theorie i = Stromdichte (A/cm2) S = Ionenart (Na, K, Cl) Um = Membranspannung
 S = Ionenart (Na, K, Cl) Um = Membranspannung.")
23
Strom-Spannungskurven für die passive Membran
lineare Näherung: iNa = gNa x (Um – ENa) Gleichgewichtspotential, Umkehrpotential, nichtlinear, monoton, Lineare Näherung für Goldman Gleichung: iges=0, ic=cm dU/dt innen gNa + Um ENa außen
 Gleichgewichtspotential, Umkehrpotential, nichtlinear, monoton, Lineare Näherung für Goldman Gleichung: iges=0, ic=cm dU/dt. innen. gNa. + Um. ENa. außen.")
24
Lineare Näherung dUm im = iNa + iK + iCl + iC iC = cm x dt dUm dt
= gNa x (Um – ENa) + gK x (Um – EK) + gCl x (Um – ECl) + cm x dUm dt Ruhezustand: Um = const. = 0 gNa ENa + gK EK + gCl ECl gNa + gK + gCl U0 = gm = gNa + gK + gCl dUm dt im = gm x (Um – U0) + cm x lin. Näherung der Goldman Gl. Kabelgleichung Nernst Gl. „Ersatzschaltbild“ für Membran
 + gK x (Um – EK) + gCl x (Um – ECl) + cm x. dUm. dt. Ruhezustand: Um = const. = 0. gNa ENa + gK EK + gCl ECl. gNa + gK + gCl. U0 = gm = gNa + gK + gCl. dUm. dt. im = gm x (Um – U0) + cm x lin. Näherung der Goldman Gl. Kabelgleichung. Nernst Gl. „Ersatzschaltbild für Membran.")
25
Kap. IV Kabelgleichung für Axon mit passiver Membran
Passive Membran gNa, gK, gCl, ENa, EK, ECl sind konstant Ändert sich die Membranspannung entlang des Axons Longitudinalströme Um = Um(x,t) IP = Strom im Axoplasma x x2 IP 2a Widerstand des Axoplasmas RP RP = rP x l/f rP = spezifischer Widerstand [W x m] f = a2p rP = rP/f = Widerstand pro Längeneinheit = 0 t = cm/gm l2 = 1/(2pa x rP x gm) = a/(2rPgm) l proportional Va
 IP = Strom im Axoplasma. x1 x2. IP. 2a. Widerstand des Axoplasmas RP. RP = rP x l/f rP = spezifischer Widerstand [W x m] f = a2p. rP = rP/f = Widerstand pro Längeneinheit. = 0. t = cm/gm l2 = 1/(2pa x rP x gm) = a/(2rPgm) l proportional Va.")
26
(4.1)
")
28
2. „Zeitklemme“: = 0 l2 x - (Um – U0) = 0 dUm dx dUm dt
Anwendungsbeispiele „Ortsklemme“: 2. „Zeitklemme“: = 0 l2 x (Um – U0) = 0 dUm dx dUm dt = 0 t x + Um = U0 dUm dt dU2m dx2 Ortsklemme Zeitklemme für x < 0 für x > 0 1: Axon O 10µ, l = 2 mm 2: Axon O 1µ, l = 0,65 mm 3: Axon O 0,1µ, l = 0,2 mm t = cm/gm ~ 5 ms (Riesenaxon)
 = 0. dUm. dx. dUm. dt. = 0 t x + Um = U0. dUm. dt. dU2m. dx2. Ortsklemme Zeitklemme. für x < 0. für x > 0. 1: Axon O 10µ, l = 2 mm. 2: Axon O 1µ, l = 0,65 mm. 3: Axon O 0,1µ, l = 0,2 mm. t = cm/gm ~ 5 ms (Riesenaxon)")
29
Wie groß ist der Gesamtstrom, den man in das Axon injizieren muss, um
am Ort x = 0 die Membranspannung Umx=0 zu erhalten? rm = 1/(2pagm) Diese Beziehung definiert den sog. Eingangswiderstand des Axons: rP = r/a2p
 Diese Beziehung definiert den sog. Eingangswiderstand des Axons: rP = r/a2p.")
30
Beispiel: Axondurchmesser = 10µ: Rinput = 3,9 MW
Axondurchmesser = 0,1µ: Rinput = 3,9 GW Werte gelten nur für Riesenaxon. Experimentell: Messung von a, Rinput und l um die Materialkonstanten rP des Axoplasmas und gm zu berechnen: Größenordnungen: gm = 2x10-4 S/cm2 rP = 30 Wcm

31
Ionale Grundlagen des Aktionspotentials
Kapitel V Ionale Grundlagen des Aktionspotentials Passiver Nervenfortsatz (Dendrit): Passive Membran, Leitfähigkeiten gNa und gK sind Konstanten. Ausbreitung des elektrischen Signals nach Kabelgleichung, räumlich und zeitlich exponentiell abfallend. Aktiver Nervenfortsatz (Axon): Aktive Membran, Aktionspotentiale, Signalleitung ohne Dekrement. Leitfähigkeiten gNa und gK hängen 1) vom momentanen Membranpotential Um 2) von der Zeit t abhängen, also Eine passive Membran vermag elektrische Signale nur nach der in Kapitel IV besprochenen Kabelgleichung fortzuleiten, also mit zeitlich und räumlich exponentiell abfallender Amplitude. Aktionspotentiale entstehen nur an aktiven Membranen. Der entscheidende Unterschied zwischen aktiven und passiven Membranen liegt in den Leitfähigkeiten von Na+ und K+. gNa und gK sind Konstanten für passive Membranen, während sie in aktiven Membranen
: Passive Membran, Leitfähigkeiten gNa und gK sind Konstanten. Ausbreitung des elektrischen Signals nach Kabelgleichung, räumlich und zeitlich exponentiell abfallend. Aktiver Nervenfortsatz (Axon): Aktive Membran, Aktionspotentiale, Signalleitung ohne Dekrement. Leitfähigkeiten gNa und gK hängen. 1) vom momentanen Membranpotential Um. 2) von der Zeit t. abhängen, also. Eine passive Membran vermag elektrische Signale nur nach der in Kapitel IV besprochenen Kabelgleichung fortzuleiten, also mit zeitlich und räumlich exponentiell abfallender Amplitude. Aktionspotentiale entstehen nur an aktiven Membranen. Der entscheidende Unterschied zwischen aktiven und passiven Membranen liegt in den Leitfähigkeiten von Na+ und K+. gNa und gK sind Konstanten für passive Membranen, während sie in aktiven Membranen.")
32
Ionale Grundlagen des Aktionspotentials
Kapitel V Ionale Grundlagen des Aktionspotentials Passiver Nervenfortsatz (Dendrit): Passive Membran, Leitfähigkeiten gNa und gK sind Konstanten. Ausbreitung des elektrischen Signals nach Kabelgleichung, räumlich und zeitlich exponentiell abfallend. Aktiver Nervenfortsatz (Axon): Aktive Membran, Aktionspotentiale, Signalleitung ohne Dekrement. Leitfähigkeiten gNa und gK hängen 1) vom momentanen Membranpotential Um 2) von der Zeit t abhängen, also gNa = gNa(Um, t) gK = gK(Um, t) Eine passive Membran vermag elektrische Signale nur nach der in Kapitel IV besprochenen Kabelgleichung fortzuleiten, also mit zeitlich und räumlich exponentiell abfallender Amplitude. Aktionspotentiale entstehen nur an aktiven Membranen. Der entscheidende Unterschied zwischen aktiven und passiven Membranen liegt in den Leitfähigkeiten von Na+ und K+. gNa und gK sind Konstanten für passive Membranen, während sie in aktiven Membranen
: Passive Membran, Leitfähigkeiten gNa und gK sind Konstanten. Ausbreitung des elektrischen Signals nach Kabelgleichung, räumlich und zeitlich exponentiell abfallend. Aktiver Nervenfortsatz (Axon): Aktive Membran, Aktionspotentiale, Signalleitung ohne Dekrement. Leitfähigkeiten gNa und gK hängen. 1) vom momentanen Membranpotential Um. 2) von der Zeit t. abhängen, also. gNa = gNa(Um, t) gK = gK(Um, t) Eine passive Membran vermag elektrische Signale nur nach der in Kapitel IV besprochenen Kabelgleichung fortzuleiten, also mit zeitlich und räumlich exponentiell abfallender Amplitude. Aktionspotentiale entstehen nur an aktiven Membranen. Der entscheidende Unterschied zwischen aktiven und passiven Membranen liegt in den Leitfähigkeiten von Na+ und K+. gNa und gK sind Konstanten für passive Membranen, während sie in aktiven Membranen.")
33
Aktionspotential: Membranpotential wird positiv nur zu erklären mit gNa+ groß.
In allen bisher untersuchten Axonen Na+-Einstrom für Aktionspotential verantwortlich. Ausnahmen: bei einigen Zellkörpern und einigen Muskelzellen ist Calcium Träger des Einwärtsstroms beim Aktionspotential. Mathematische Beschreibung der Vorgänge am aktiven Axon: Phänomenologie, physikalische Ableitung wie beim Ruhepotential nicht möglich. Voraussetzung für quantitative Analyse der Membranströme: Um sprunghaft verändern und dann konstant halten („Spannungsklemme“, voltage clamp). iC = 0 ca. 10 µsec nach Spannungssprung (iC = kapazitiver Strom vernachlässigbar)
. iC = 0 ca. 10 µsec nach Spannungssprung (iC = kapazitiver Strom vernachlässigbar)")
34
Niederohmiges Netzgerät mit Strom- und Spannungsanzeige
Im Prinzip: Niederohmiges Netzgerät mit Strom- und Spannungsanzeige (und Oszilloskop) Problem: Polarisationsspannung an Grenzfläche zwischen Metall und Elektrolyt, variabel, hängt von Strom ab, tatsächliche Membranspannung unbekannt. Lösung: x1 Verstärker: hoher Eingangswiderstand, nahezu stromlose Messung, keine Polarisationsspannung an oberer Elektrode. Polarisationsspannung an unterer Elektrode wird durch Differenzverstärker kompensiert. „Two-electrode voltage clamp“ (TEVC)
 Problem: Polarisationsspannung an Grenzfläche zwischen Metall und Elektrolyt, variabel, hängt von Strom ab, tatsächliche Membranspannung unbekannt. Lösung: x1 Verstärker: hoher Eingangswiderstand, nahezu stromlose Messung, keine. Polarisationsspannung an oberer Elektrode. Polarisationsspannung an unterer. Elektrode wird durch Differenzverstärker kompensiert. „Two-electrode voltage clamp (TEVC)")
35
Ersatzschaltbild mit variablen Leitfähigkeiten für Natrium und Kalium:
Wie kann man die Ionenströme für Natrium und Kalium trennen? 2 Verfahren: 1) Veränderung der Ionenkonzentrationen, so dass (Um – ENa) bzw. (Um – EK) null wird
 Veränderung der Ionenkonzentrationen, so dass. (Um – ENa) bzw. (Um – EK) null wird.")
36
Beispiel: normale Ionenkonzentrationen: Sprung auf -9 mV Kurve A
Änderung von Na+außen so dass ENa = - 9 mV iNa = 0 Kurve B = iK Differenz aus Kurve B und Kurve A: Kurve C = iNa Voraussetzung: Ionenfluss von Natrium unabhängig von Kaliumkonzentration (und umgekehrt): „Unabhängigkeitsprinzip“
: „Unabhängigkeitsprinzip")
37
2) Pharmakologische Trennung der Ionenströme:
Tetrodotoxin (TTX, aus Ovarien des Pufferfisch) und Saxitoxin (STX) blockieren selektiv die Natriumkanäle (von außen) Tetraethylamonium (TEA) blockiert selektiv die Kaliumkanäle (von innen) Antworten auf Spannungssprünge verschiedener Größe:
 und Saxitoxin (STX) blockieren. selektiv die Natriumkanäle (von außen) Tetraethylamonium (TEA) blockiert selektiv die Kaliumkanäle (von innen) Antworten auf Spannungssprünge verschiedener Größe:")
38
Diese Versuche zeigen:
iNa : Bei Depolarisation schneller Einwärtsstrom, der schnell abklingt, obwohl die Depolarisation anhält. iK : Bei Depolarisation verzögerter Auswärtsstrom, der bei anhaltender Depolarisation anhält. „früher Spitzenstrom“ IP = iNa „stationärer Strom“ ISS = IK Werden Na-Kanäle durch TTX blockiert, so bleibt ein kleiner früher Spitzenstrom übrig, der dem Ohm-schen Gesetz folgt: „Leckstrom“ mehrerer Ionenarten = IL Nulldurchgang: EL Qualitative Interpretation: Aktionspotential (negativer Widerstand, „alles oder nichts“)
")
39
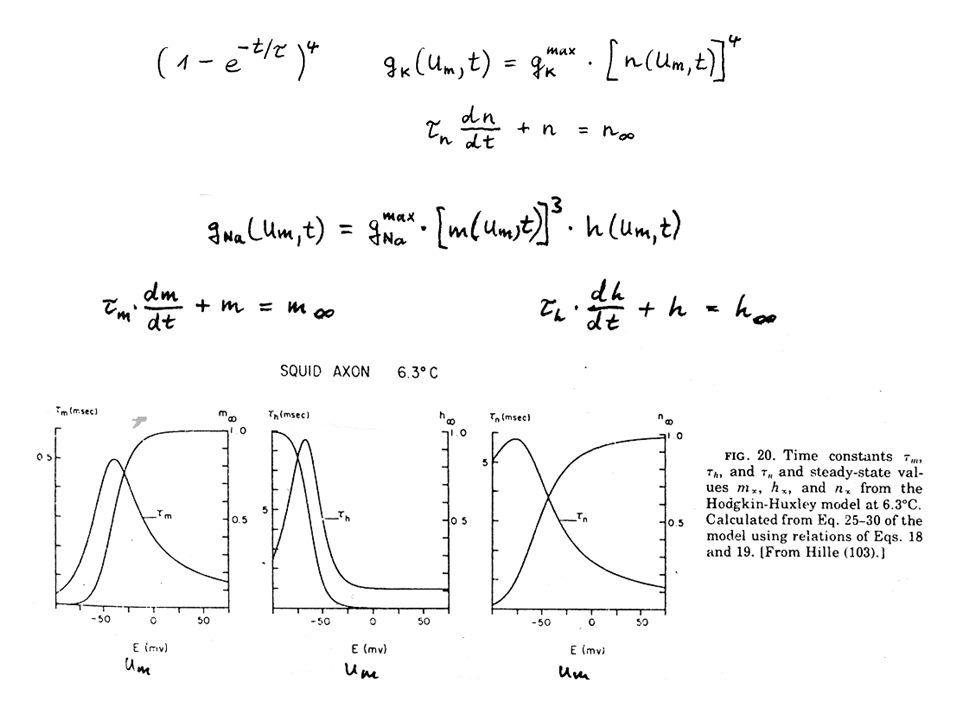
Hodgkin-Huxlex Gleichungen
Können die unter Spannungklemme als Funktion der Zeit gemessenen Ströme das Verhalten des Axons unter Normalbedingungen vorhersagen? iK = gKx(Um – EK) iNa = gNax(Um – ENa) Problem: Stromverlauf bei jeder Spannung anders gK = gK(Um, t) gNa = gNa(Um, t) Beschreibung nur durch Differentialgleichungen möglich. Im Prinzip viele Lösungen möglich, Suche nach der einfachsten.
 iNa = gNax(Um – ENa) Problem: Stromverlauf bei jeder Spannung anders gK = gK(Um, t) gNa = gNa(Um, t) Beschreibung nur durch Differentialgleichungen möglich. Im Prinzip viele Lösungen möglich, Suche nach der einfachsten.")
41
Lösung der Hodgin-Huxley Gleichungen
a) Ortsklemme: gute Übereinstimmung mit Experiment
 Ortsklemme: gute Übereinstimmung mit Experiment.")
42
b) Reizfortleitung
 Reizfortleitung")
43
4 Phasen erläutern

Ähnliche Präsentationen