Präsentation herunterladen
Die Präsentation wird geladen. Bitte warten

1
Steuerung des Zellzyklus
nach: Alberts Kapitel 17

2
Der Zellzyklus im Mikroskop

3
Die 4 Phasen des Zellzyklus

4
Steuerung des Zellzyklus
Modelle zum Studium des Zellzyklus-Steuerungssystems Die Komponenten des Zellzyklus-Steuerungssystems Intrazelluläre Kontrolle der Ereignisse im Zellzyklus

5
Die Genetik der Steuerung des Zellzyklus lässt sich gut in (haploiden) Hefen untersuchen. Vergleich von Spalthefe und Bäckerhefe („Knospende Hefe“)
.")
6
Die Genetik der Steuerung des Zellzyklus lässt sich gut in (haploiden) Hefen untersuchen. Vergleich von Spalthefe und Bäckerhefe („Knospende Hefe“)
.")
7
Temperatur-sensitive cdc-Mutanten (cdc: cell-division-cycle genes)
Haploide Hefe cdc-Mutanten teilen sich normal bei tiefen Temperaturen Bei höheren Temperaturen funktioniert das mutierte Genprodukt nicht mehr, die Zellen bleiben in einem bestimmten Zellstadium stehen.

8
Temperatur-sensitive cdc-Mutanten (cdc: cell-division-cycle genes)
Haploide Hefe cdc-Mutanten teilen sich normal bei tiefen Temperaturen Bei höheren Temperaturen funktioniert das mutierte Genprodukt nicht mehr, die Zellen bleiben in einem bestimmten Zellstadium stehen.

9
Die riesige Eizelle lässt sich leicht handhaben
Die Biochemie der Steuerung des Zellzyklus lässt sich gut in Xenopus Eiern untersuchen. Die riesige Eizelle lässt sich leicht handhaben

10
Oozytenwachstum und mitotische Teilungen

11
Studium des Zellzyklus im zellfreien System
Oozyten werden durch Zentrifugation aufgebrochen. Zum Zytoplasma werden Kerne von Spermien zugegeben. Die Kerne führen unter diesen Bedingungen mehrere mitotische Teilungen durch.

12
Kulturzellen von Vertebraten

13
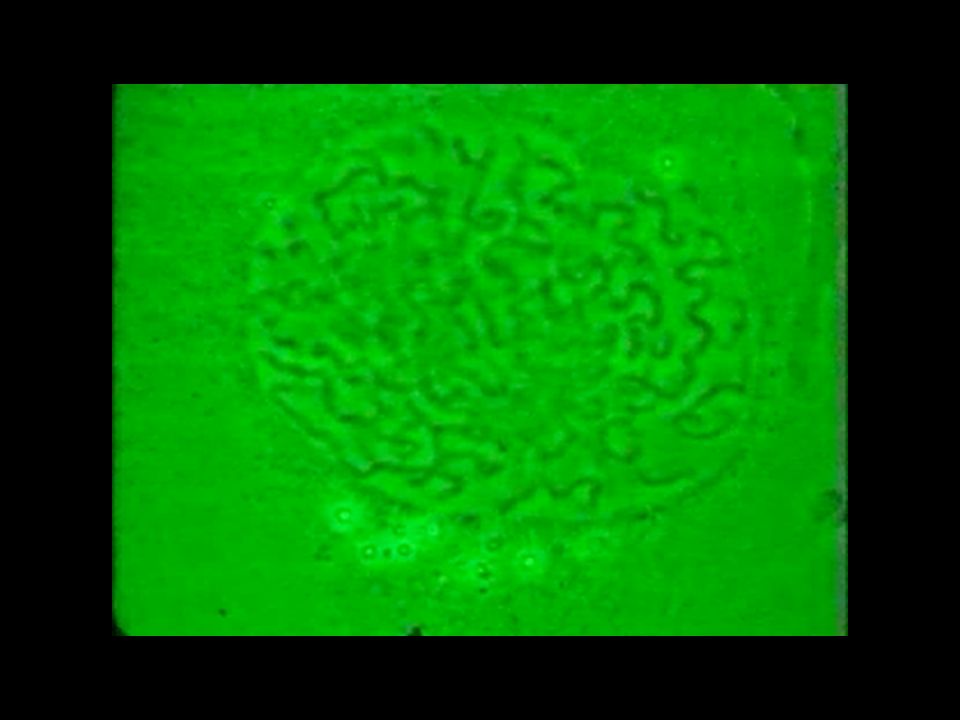
Markierung von S-Phase Zellen (im Darm eines Zebrafisches)
durch Zugabe von BrdU (Bromo-Deoxyuridin) einem Thymidin-Analogen , das in neu synthetisierte DNA eingebaut wird und immunologisch mit grün fluoreszierenden Antikörpern nachgewiesen werden kann.
 einem Thymidin-Analogen , das in neu synthetisierte DNA eingebaut wird und immunologisch mit grün fluoreszierenden Antikörpern nachgewiesen werden kann.")
14
DNA-Gehalt gemessen im Flow-Zytometer (FACS; fluorescence activated cell sorter)
Fluoreszenzfarbstoff bindet an DNA; Auftrennung der Zellen nach DNA-Gehalt In der Population befinden sich mehr Zellen in der G1-Phase als in S- und G2-Phase.

15
Steuerung des Zellzyklus
Modelle zum Studium des Zellzyklus-Steuerungssystems Die Komponenten des Zellzyklus-Steuerungssystems Intrazelluläre Kontrolle der Ereignisse im Zellzyklus

16
Das Zellzyklus-Steuerungssystem
Anforderungen an das Steuerungssystem: Eine Schaltuhr, die die Ereignisse in der richtigen Reihenfolge und nur einmal schaltet. Die Schaltung soll binär erfolgen, keine halben Sachen. Das System muss zuverlässig funktionieren und durch Backup-Systeme abgesichert sein. Das System muss für spezielle Ansprüche modifiziert werden können. Bei einem „einfachen“ Zellzyklus; z.B. in Embryonen erfolgt die Steuerung wie bei der Schaltuhr einer Waschmaschine. In den meisten Fällen erfolgt aber Rückkopplung bei bestimmten Checkpoints. Ein ölöl

17
Rückkopplung bei Checkpoints
Bei den Checkpoints kann der Zellzyklus gestoppt werden, bis das Rückkopplungssignal erfolgt. Signale zur Regulation des Zekllzyklus können auch von anderen Zellen kommen. Die Signale sind oft negativ, d.h. Abwesenheit eines bestimmten Signals löst den Schalter aus. (Beispiel: Start der Anaphase)
")
18
Cyklin-abhängige Kinasen Cdks; cyclin-dependent kinases
Cdk ist eine Proteinkinase, die durch Cyclin aktiviert wird.

19
Protein Phosphorylierung

20
Das Zellzyklus-Steuerungssystem
(vereinfachte Darstellung) Cdks verbinden sich mit verschiedenen Cyclinen um einen Prozess einzuschalten. Die Aktivität wird durch Cyclin-Abbau beendet.
 Cdks verbinden sich mit verschiedenen Cyclinen um einen Prozess einzuschalten. Die Aktivität wird durch Cyclin-Abbau beendet.")
21
Die strukturelle Basis der Cdk-Aktivierung durch Cyclin
Bindung von Cyclin bewirkt Verschiebung des T-Loop. Phosphorylierung bewirkt weitere Veränderung des T-Loop und erhöht Aktivität. Die aktive Stelle ist durch den T-Loop abgedeckt.

22
Weitere Phosphorylierungen inhibieren Cdk-Aktivität
Wee1 Kinase kann 2 zusätzliche Phosphatgruppen anbringen (nur eine gezeichnet); bewirkt Inaktivierung der Cdk. Cdc 25 (cell division cycle) kann diese wieder entfernen und bewirkt Aktivierung.
; bewirkt Inaktivierung der Cdk. Cdc 25 (cell division cycle) kann diese wieder entfernen. und bewirkt Aktivierung.")
23
CKIs inhibieren den Cyclin-Cdk-Komplex
CKI: Cdk Inhibitor Protein. Wichtige Funktion bei der Kontrolle von G1- und S-Phase.

24
Cyclin dependent Kinases (CDK)
-transferrieren Phosphat-Gruppen von ATP auf Serin- und Threonin-Seitenketten des Substrats. -Im inaktiven Zustand ist aktive Stelle durch T-loop abgedeckt -CDKs werden durch Cykline aktiviert. -Substrat bindet an spezifische Stelle. -CDK Inhibitors (CDKI)
")
25
Steuerung der Proteolyse von Cyclin durch APC (Anaphase Promoting Complex)
(Ubiquitin ist ein kleines Protein mit 76 Aminosäuren; es kann am C-Terminus an Proteine gebunden werden, die dann in den Proteasomen abgebaut werden.)
")
26
Steuerung der Proteolyse von CKI durch SCF
Ein aktivierter Proteinkomplex (SCF) erkennt bestimmte Aminosäurensequenzen und bindet Ubiquitin daran, um es für den Abbau in den Proteasomen zu markieren.
 erkennt bestimmte Aminosäurensequenzen und bindet Ubiquitin daran, um es für den Abbau in den Proteasomen zu markieren.")
27
Zusammenfassung Die Ereignisse des Zellzyklus sind durch ein unabhängiges Kontrollsystem gesteuert. Das Kontrollsystem kann durch Rückkopplung den Zellzyklus nötigenfalls zu stoppen. Die Komponenten sind cyklin-abhängige Protein-Kinasen (CdKs), deren Aktivität durch Cyklin gesteuert wird. Oszillationen in der Aktivität der verschiedenen Cyklin-Komplexe steuert die Inititiation der Ereignisse. Verschiedene Kontrollmechanismen beeinflussen die aktivität der Cdks.
, deren Aktivität durch Cyklin gesteuert wird. Oszillationen in der Aktivität der verschiedenen Cyklin-Komplexe steuert die Inititiation der Ereignisse. Verschiedene Kontrollmechanismen beeinflussen die aktivität der Cdks.")
28
Steuerung des Zellzyklus
Modelle zum Studium des Zellzyklus-Steuerungssystems Die Komponenten des Zellzyklus-Steuerungssystems Intrazelluläre Kontrolle der Ereignisse im Zellzyklus

29
Übersicht über das Zellzyklus-Kontrollsystem
Das Herz der Steuerung sind die verschiedenen Cyclin-Cdk-Komplexe. Deren Aktivität kann an den Checkpoints durch verschiedene Proteine gehemmt werden. Diese Proteine und Hemmechanismen sind nicht in allen Zellen vorhanden, viele kommen z. B. in sich schnell teilenden embryonalen Zellen nicht vor.

30
Eintritt in Mitose

31
Die Aktivierung von M-Cdks zu Beginn der Mitose
Die Konzentration von M-Cyklin steigt in G2 durch stärkere Gen-Transkription. Positive Rückkoppelung führt zu sprunghaftem Anstieg von aktiver M-Cdk; dies fördert einen binären alles-oder-nichts-Effekt.

32
17-18 Weitere Phosphorylierungen inhibieren Cdk-Aktivität
Wee1 Kinase kann 2 zusätzliche Phosphatgruppen anbringen (nur eine gezeichnet); bewirkt Inaktivierung der Cdk. Cdc 25 (cell division cycle) kann diese wieder entfernen und bewirkt Aktivierung.
; bewirkt Inaktivierung der Cdk. Cdc 25 (cell division cycle) kann diese wieder entfernen. und bewirkt Aktivierung.")
33
Funktionen von M-Cdks M-Cdks Eintritt in Mitose:
starten die Chromosomenkondensation. phosphorylieren die Lamine (führt zu deren Aufbrechen und so zum Abbau der Kernhülle). induzieren den Aufbau der Spindel durch Erhöhen der Mikrotubuli-Instabilität. Eintritt in Anaphase: induzieren (indirekt) die Anaphase. starten die Organisation des Kontraktilen Rings. starten die Reorganisation von Golgi und ER.
. induzieren den Aufbau der Spindel durch Erhöhen der Mikrotubuli-Instabilität. Eintritt in Anaphase: induzieren (indirekt) die Anaphase. starten die Organisation des Kontraktilen Rings. starten die Reorganisation von Golgi und ER.")
34
Ab-und Aufbau der nukleären Lamina

35
Eintritt in Anaphase:

36
APC triggert die Teilung der Chromosomen in Chromatiden zwischen Meta- und Anaphase
Cdc20-Transkription steigt vor der Mitose an. Phosphorylierung von APC fördert Bindung mit Cdc20. APC: Anaphase Promoting Complex Cdc: Cell division cycle protein

37
APC triggert die Teilung der Chromosomen in Chromatiden zwischen Meta- und Anaphase

38
Mad2 Protein findet sich an Kinetochoren, die noch nicht an beide Pole gebunden sind

39
Inhibiert, wenn einChromosom noch nicht beidseitig am Spindelapparat hängt

41
Übergang von M-Phase in G1
Die durch M-Cdk mittels Phophorylierung iniziierten Prozesse werden durch Inhibition von M-Cyklinen und durch Phosphatasen (durch Dephosphorylierung z. B. der Lamine) abgeschlossen.
 abgeschlossen.")
42
Übergang von G1 zu S-Phase

43
17-60Cdk-Inhibition zum Start der G1-Phase
M-Cyclin wird durch den Cdc20-APC Komplex ubiquitiniliert und abgebaut. (Dies bewirkt aber auch Reduktion der Cdc20-APC Aktivität, da diese durch M-Cdk stimuliert wird). Sofortiger Wiederanstieg von M-Cyclin verhindert G1-Phase.
. Sofortiger Wiederanstieg von M-Cyclin verhindert G1-Phase.")
44
17-60Cdk-Inhibition zum Start der G1-Phase
M-Cyclin wird durch den Cdc20-APC Komplex ubiquitiniliert und abgebaut. (Dies bewirkt aber auch Reduktion der Cdc20-APC Aktivität, da diese durch M-Cdk stimuliert wird). Sofortiger Wiederanstieg von M-Cyclin verhindert G1-Phase. M-Cyclin wird durch Hct1-APC niedrig gehalten; dies verhindert Einstieg in S-Phase und verlängert die G1-Phase. (Hct1 ist verwandt mit Cdc 20 wird aber durch M-Cdk nicht aktiviert sondern gehemmt)
. Sofortiger Wiederanstieg von M-Cyclin verhindert G1-Phase. M-Cyclin wird durch Hct1-APC niedrig gehalten; dies verhindert Einstieg in S-Phase und verlängert die G1-Phase. (Hct1 ist verwandt mit Cdc 20 wird aber durch M-Cdk nicht aktiviert sondern gehemmt)")
45
ENDE

Ähnliche Präsentationen



>")















