Präsentation herunterladen
Die Präsentation wird geladen. Bitte warten

1
Physio 1 WS (2st.) Physio 2 SS (1st.)
PHYSIOLOGIE Physio 1 WS (2st.) Physio 2 SS (1st.)
 Physio 2 SS (1st.)")
2
Inhalte VO Physiologie 1
Grundlagen der Physiologie der Muskelkontraktion Grundlagen der elektrischen Erregung (RP, AP) Erregungsübertragung von Zelle zu Zelle Molekulare Mechanismen der Kontraktion Mechanismen der Kraftregulation, Physiologe des Kraftrainings Grundlagen der Endokrinologie Aufbau und Funktion der Hormone Sport und hormonelle Steuerung
 Erregungsübertragung von Zelle zu Zelle. Molekulare Mechanismen der Kontraktion. Mechanismen der Kraftregulation, Physiologe des Kraftrainings. Grundlagen der Endokrinologie. Aufbau und Funktion der Hormone. Sport und hormonelle Steuerung.")
3
Inhalte VO Physiologie 1
Energiebereitstellung Aerober/Anaerober Stoffwechsel Fettstoffwechsel Body Composition Übergewicht/Adipositas Sport und Körperfettabbau Regulationsaspekte des Körpergewichts (Anorexia Athletica)
")
4
Empfehlenswerte Literatur
Levick, JR. Physiologie des Herz-Kreislauf-Systems Schmidt, Thews: Physiologie des Menschen Thews, Mutschler, Vaupel: Anatomie, Physiologie, Pathophysiologie des Menschen De Marées, Horst: Sportphysiologie Kandel, ER., Schwartz, JH., Jessell, TM.(eds) Neurowissenschaften. Bray, GA., Bouchard C., James, WPT.: Handbook of Obesity Roche, Heymsfield, Lohman: Human Body Composition Davies, Cole: Body Composition Techniques in Health and Disease Logue, AW. Die Psychologie des Essens und Trinkens American Journal of Physiology [ American Journal of Clinical Nutrition [ Society for Endocrinology-Electronic Journals [ Medicine and Science in Sports and Exercise [
 Neurowissenschaften. Bray, GA., Bouchard C., James, WPT.: Handbook of Obesity. Roche, Heymsfield, Lohman: Human Body Composition. Davies, Cole: Body Composition Techniques in Health and Disease. Logue, AW. Die Psychologie des Essens und Trinkens. American Journal of Physiology [ American Journal of Clinical Nutrition [ Society for Endocrinology-Electronic Journals [ Medicine and Science in Sports and Exercise [")
5
!!! (hoffentlich) POSITIVE NOTE !!!
Prüfungsmodalitäten Schriftlich (mündlich nur in besonderen Fällen nach Vereinbarung) Termin am Ende des Semesters Termin 4-6 Wochen später Termin Mitte des folgenden Semesters !!! (hoffentlich) POSITIVE NOTE !!!
 Termin am Ende des Semesters. Termin 4-6 Wochen später. Termin Mitte des folgenden Semesters. !!! (hoffentlich) POSITIVE NOTE !!!")
6
Physiologie des Erregungsaufbaus und - überleitung
Nervenzellen und Muskelzellen besitzen ein negatives Ruhemembranpotential (~ - 55 bis -100mV) Ruhepotential: weitgehend ein K+ Diffusionspotential Extrazellulärraum Ionen K+ Intrazellulärraum Ionen Na+
 Ruhepotential: weitgehend ein K+ Diffusionspotential. Extrazellulärraum. Ionen. K+ Intrazellulärraum. Ionen. Na+")
7
Physiologie des Erregungsaufbaus und - überleitung
Stoffaustausch der Zelle die verschiedenen Räume der Zelle (und die Zelle) stehen mit ihrer Umgebung in ständigem Stoffaustausch Der einfachste Stoffaustausch ist die DIFFUSION (Ausgleich von Konzentrationsunterschieden) Verschiedene Stoffe (u.a. H2O, gelöste Gase, lipidlösliche Stoffe, Ethanol, Harnstoff) können frei durch die Zellmembran diffundieren - für geladene Moleküle (Ionen) ist die Membran de facto impermeabel; Ionen passieren die Membran durch Kanäle in Proteinmolekülen
 stehen mit ihrer Umgebung in ständigem Stoffaustausch. Der einfachste Stoffaustausch ist die DIFFUSION. (Ausgleich von Konzentrationsunterschieden) Verschiedene Stoffe (u.a. H2O, gelöste Gase, lipidlösliche Stoffe, Ethanol, Harnstoff) können frei durch die Zellmembran diffundieren - für geladene Moleküle (Ionen) ist die Membran de facto impermeabel; Ionen passieren die Membran durch Kanäle in Proteinmolekülen.")
8
Physiologie des Erregungsaufbaus und - überleitung
Poren bzw. Kanäle gebildet von in den Membranen eingelagerten Transportproteinen; derartige Membranproteine werden durch einen kleinen Kanal durchzogen - die diffundierenden Substanzen bewegen sich durch den Kanal entsprechend der Konzentrationsgradienten und unter dem Einfluss des Membranpotentials Membrankanäle sind relativ selektiv hinsichtlich der durchfließenden Moleküle zB. Kalium-, Natrium-, Kalziumkanäle

9
Physiologie des Erregungsaufbaus und - überleitung
Aktiver Transport aufgrund der Diffusion würden sich die IZ-[Na+] und [K+] den EZ-Konzentrationen angleichen; Membranproteine transportieren Ionen über die Membran dem Konzentrations- bzw. elektr. Gradienten entgegen, wobei Stoffwechselenergie verbraucht wird Der wichtigste aktive Transportprozeß ist die Na-K-Pumpe, die Na+ aus der Zelle, und K+ in die Zelle bringt damit wird die niedrige IZ-[Na+] und hohe [K+] sichergestellt ~ 1/3 des Energieverbrauchs der Zelle wird für die Na-K-Pumpe verbraucht (bis 70%)
")
10
Physiologie des Erregungsaufbaus und - überleitung
Na-K-Pumpe ist eine ATPase, spaltet an der Innenseite der Membran ATP in ADP und Phosphat auf - dabei werden 3 Na+ aus der Zelle und gleichzeitig 2 K+ in die Zelle gepumpt Na-K-Pumpe ist elektrogen (sie treibt einen elektrischen Strom über die Membran aus der Zelle) Die Na-K-Pumpe ist a.) stark temperatur- und energieabhängig b.) und die Pumprate erreicht einen Sättigungswert Ca-Pumpe [diese pumpt Ca2+ Ionen aus der Zelle und ist somit an der niedrigen IZ [Ca2+] beteiligt]
 Die Na-K-Pumpe ist. a.) stark temperatur- und energieabhängig. b.) und die Pumprate erreicht einen Sättigungswert. Ca-Pumpe [diese pumpt Ca2+ Ionen aus der Zelle und ist somit an der niedrigen IZ [Ca2+] beteiligt]")
11
Entstehung und Fortleitung von Aktionspotentialen
Funktion von Nervenzellen: Informationsaufnahme, Informationsweiterleitung, und Informationsintegration damit werden die Funktionen anderer Zellen gesteuert Axon Soma, Perikaryon Axonhügel Dendriten Kern

12
Entstehung und Fortleitung von Aktionspotentialen
Soma: Stoffwechselzentrum Dendrit: verzweigen sich, und dienen der Informations (Signal) aufnahme von anderen Zellen Axon: röhrenförmiger Fortsatz, wichtigste Leitungseinheit des Neurons (leitet Signale bis zu 2m) Jene Signale, die über das Axon geleitet werden sind Aktionspotentiale (schnelle, kurzlebige Alles- od-Nichts Nervenimpulse; Amplitude ~ 100mV, Dauer ~ 1ms, v ~ 1-100m/s) die am Axonhügel entstehen
 aufnahme von anderen Zellen. Axon: röhrenförmiger Fortsatz, wichtigste Leitungseinheit des Neurons (leitet Signale bis zu 2m) Jene Signale, die über das Axon geleitet werden sind Aktionspotentiale (schnelle, kurzlebige Alles- od-Nichts Nervenimpulse; Amplitude ~ 100mV, Dauer ~ 1ms, v ~ 1-100m/s) die am Axonhügel entstehen.")
13
Entstehung und Fortleitung von Aktionspotentialen
Um die rasche Fortleitung der AP-Signale zu gewährleisten, besitzen die großen Axone eine lipidhaltige Isolierung (Myelin). Eine derartige Myelinscheide ist von nichtisolierten Stellen unterbrochen (Ranvier-Schnürringen) - an diesen Stellen wird das AP ‚regeneriert‘ Das AP entsteht aus einem verstärkten Fluß von Ionen durch die Membran
. Eine derartige Myelinscheide ist von nichtisolierten Stellen unterbrochen (Ranvier-Schnürringen) - an diesen Stellen wird das AP ‚regeneriert‘ Das AP entsteht aus einem verstärkten Fluß von Ionen durch die Membran.")
15
- + EZF Na+ K+ K+ Na+ IZF Diffusions potential K+
[K+,Na+,..]EZF ~ [K+, Na+, ...]IZF

16
[K+,Na+,..]EZF ~ [K+, Na+, ...]IZF
Gleichgewichtskonz. Gleichgewichtspot. Ruhemembranpotential ~ mV (je nach Zelltyp) EZF Ungleiche Ionen- verteilung IZF
![[K+,Na+,..]EZF ~ [K+, Na+, ...]IZF](http://slideplayer.org/slide/874998/2/images/16/%5BK%2B%2CNa%2B%2C..%5DEZF+%7E+%5BK%2B%2C+Na%2B%2C+...%5DIZF.jpg "Gleichgewichtskonz. Gleichgewichtspot. Ruhemembranpotential. ~ mV (je nach Zelltyp) EZF. Ungleiche Ionen- verteilung. IZF.")
17
Aktiver Transport Na+ EZF K+ Na+ IZF K+ [K+] [Na+]
![Aktiver Transport Na+ EZF K+ Na+ IZF K+ [K+] [Na+]](http://slideplayer.org/slide/874998/2/images/17/Aktiver+Transport+Na%2B+EZF+K%2B+Na%2B+IZF+K%2B+%5BK%2B%5D+%5BNa%2B%5D.jpg "Aktiver Transport Na+ EZF K+ Na+ IZF K+ [K+] [Na+]")
18
Depolarisationsphase
Reiz AP (-) Ruhemembranpot. 0 mV Depolarisation (Schwellenpot.) IZF Na+ Na+ K+ Depolarisationsphase des AP; + mV „Overshoot“ EZF K+
 Ruhemembranpot. 0 mV. Depolarisation. (Schwellenpot.) IZF. Na+ Na+ K+ Depolarisationsphase. des AP; + mV. „Overshoot EZF. K+")
19
+/- 0 Ruhepot. Abs. Refraktärperiode Rel. Refraktärperiode Potential
Depolarisation Repolarisation Potential Schwelle Ruhepot. Zeit Abs. Refraktärperiode Rel. Refraktärperiode

20
Entstehung und Fortleitung von Aktionspotentialen
Fazit 1. Eine Depolarisation der Membran verursacht ein plötzliches Öffnen der Na+ Kanäle (Ansteigen der Na-Leitfähigkeit) und führt zu einem Na+ Einwärtsstrom. 2. Dieser Strom bewirkt eine stärkere Depolarisation und öffnet noch mehr Na+ Kanäle Verstärkung des Na+ Einwärtsstrom; durch diese positive Rückkoppelung entsteht ein AP
 und führt zu einem Na+ Einwärtsstrom. 2. Dieser Strom bewirkt eine stärkere Depolarisation und öffnet noch mehr Na+ Kanäle Verstärkung des Na+ Einwärtsstrom; durch diese positive Rückkoppelung entsteht ein AP.")
21
Entstehung und Fortleitung von Aktionspotentialen
3. Die dadurch hervorgerufene Depolarisation begrenzt die Dauer des AP: a.) nach und nach werden die Na+ Kanäle inaktiviert (damit sinkt die Na-Leitfähigkeit) b.) die K+ Kanäle werden geöffnet (dadurch steigt die K-Leitfähigkeit) dem Na+ Einstrom folgt ein K+ Ausstrom, der die Membran wieder repolarisiert 4. Dem AP folgt eine Phase der verringerten Erregbarkeit (Abs. RZ: Zelle kann nicht erregt werden; Rel. RZ: AP kann erneut ausgelöst werden, wenn die Reize stärker sind als normal notwendig)
 nach und nach werden die Na+ Kanäle inaktiviert (damit sinkt die Na-Leitfähigkeit) b.) die K+ Kanäle werden geöffnet (dadurch steigt die K-Leitfähigkeit) dem Na+ Einstrom folgt ein K+ Ausstrom, der die Membran wieder repolarisiert. 4. Dem AP folgt eine Phase der verringerten Erregbarkeit (Abs. RZ: Zelle kann nicht erregt werden; Rel. RZ: AP kann erneut ausgelöst werden, wenn die Reize stärker sind als normal notwendig)")
22
Entstehung und Fortleitung von Aktionspotentialen
5. Das AP zeigt ein charakteristisches Alles-oder-Nichts-Verhalten [unterschwellige Reize lösen kein AP aus] Schwellenwert: jener Punkt, an dem sich der ionengetragene Nettostrom (der sich aus dem einwärtsgerichteten Na-Einstrom, und den beiden auswärtsgerichteten Strömen, dem K-Strom + dem sog. Leckstrom zusammensetzt) gerade von einem auswärtsgerichteten zu einem einwärts gerichteten umkehrt
 gerade von einem auswärtsgerichteten zu einem einwärts gerichteten umkehrt.")
23
Signalübertragung zwischen den Zellen
Der Ort der Übertragung zwischen kommunizierenden Zellen wird als Synapse bezeichnet Synapse: besteht aus 3 Elementen; präsynaptischen Endigung postsynaptischen Zelle Kontaktzone (je nach Organisation der Kontaktzone werden Synapsen in elektrische und chemische Synapsen unterteilt) Kontaktzone; synaptischer Spalt postsynaptische Zelle präsynaptische Endigung
 Kontaktzone; synaptischer Spalt. postsynaptische Zelle. präsynaptische Endigung.")
24
Signalübertragung zwischen den Zellen
Elektrische Synapsen: ermöglichen eine verzögerungsfreie Signalübertragung, treten gewöhnlich zwischen einer großen präsynaptischen Nervenfaser und einem kleinen postsynaptischen Neuron auf, haben einen schmalen synaptischen Spalt, der Strom der durch ein AP im präsynaptischen Neuron erzeugt wird, fließt durch Verbindungskanäle [Gap Junctions] direkt in die postsynaptische Zelle; elektrische Synapsen leiten in beide Richtungen (bidirektional)
")
25
Signalübertragung zwischen den Zellen
Chemische Synapsen: haben einen breiten synaptischen Spalt, eine Änderung des Membranpotentials der präsynaptischen Zelle führt zu einer Freisetzung von chemischen Botenstoffen ((Neuro-)Transmitter) aus der präsynaptischen Endigung. Der chemische Transmitter diffundiert durch den synaptischen Spalt, bindet an Rezeptormoleküle der postsynaptischen Zelle und öffnet dadurch Ionenkanäle, durch die Strom fließt (synaptische Verzögerung) Diese Form der chemischen Übertragung kann Signale verstärken
Transmitter) aus der präsynaptischen Endigung. Der chemische Transmitter diffundiert durch den synaptischen Spalt, bindet an Rezeptormoleküle der postsynaptischen Zelle und öffnet dadurch Ionenkanäle, durch die Strom fließt (synaptische Verzögerung) Diese Form der chemischen Übertragung kann Signale verstärken.")
27
Signalübertragung zwischen den Zellen
2 große Gruppen von Transmitterrezeptoren 1. Rezeptor ist Teil des Ionenkanals (kontrolliert den Kanal direkt) 2. wirkt indirekt auf den Ionenkanal, und aktiviert second messenger und andere molekulare Reaktionen innerhalb der postsynaptischen Zelle Beide Rezeptorgruppen können eine Erregung (exzitatorisch) oder Hemmung (inhibitorisch) vermitteln; dies hängt von der Struktur des Rezeptors ab, nicht vom Transmitter
 2. wirkt indirekt auf den Ionenkanal, und aktiviert second messenger und andere molekulare Reaktionen innerhalb der postsynaptischen Zelle. Beide Rezeptorgruppen können eine Erregung (exzitatorisch) oder Hemmung (inhibitorisch) vermitteln; dies hängt von der Struktur des Rezeptors ab, nicht vom Transmitter.")
28
Signalübertragung zwischen den Zellen
Wesentliche Merkmale von Transmitter-Rezeptoren 1. Membrandurchspannende Proteine, wobei die Region, die dem Außenmedium der Zelle zugekehrt ist, den aus der präsynaptischen Zelle freigesetzten Transmitter erkennt 2. Sie fungieren innerhalb der Zelle als Effektoren, indem sie das Öffnen und Schließen der Ionenkanäle steuern

29
Signalübertragung zwischen den Zellen
Neuromuskuläre Synapse Synapse zwischen Motoneuron und Skelettmuskelfaser Funktioneller Aufbau: Normalerweise wird eine einzelne Skelettmuskelfaser nur von einem einzigen motorischen Axon innerviert. Der von den Endigungen freigesetzte Neurotransmitter ist Acetylcholin (ACh), der Rezeptor auf der Muskelseite ist der nikotinische ACh-Rezeptor [2 Typen: nicotinische, muscarinische]
, der Rezeptor auf der Muskelseite ist der nikotinische ACh-Rezeptor [2 Typen: nicotinische, muscarinische]")
30
Neuromuskuläre Synapse
Funktioneller Aufbau: Das Axon des Motoneurons innerviert die motorische Endplatte [= spezialisierte Region der Muskelmembran] In der Nähe der Endplatte verliert das Axon seine Myelinscheide und zweigt sich in mehrere dünne Äste auf (deren Endigungen sind die synapt. Endknöpfchen); aus den Endigungen wird der Transmitter freigesetzt Das exzitatorische postsynaptische Potential (hervorgerufen von einem einwärtsgerichteten Strom) im Muskel wird als ‚Endplattenpotential‘ bezeichnet [hohe Amplitude!]
; aus den Endigungen wird der Transmitter freigesetzt. Das exzitatorische postsynaptische Potential (hervorgerufen von einem einwärtsgerichteten Strom) im Muskel wird als ‚Endplattenpotential‘ bezeichnet [hohe Amplitude!]")
31
Neuromuskuläre Synapse
Funktioneller Aufbau: Die Anzahl der ACh-aktivierten Kanäle hängt von der ausgeschütteten ACh-Menge ab (durch Depolarisation der postsynaptischen Zelle aktivieren die transmittergesteuerten Kanäle spannungsgesteuerte Na+ Kanäle in der Endplattenregion) Mit zunehmender Depolarisation der postsynaptischen Zelle werden immer mehr Na+ Kanäle aktiviert – der resultierende Na+ Einstrom führt dann zur Auslösung eines AP‘s
 Mit zunehmender Depolarisation der postsynaptischen Zelle werden immer mehr Na+ Kanäle aktiviert – der resultierende Na+ Einstrom führt dann zur Auslösung eines AP‘s.")
33
Molekulare Mechanismen der
Muskelkontraktion kontraktile Proteine….. 1g Skelettmuskel enthält ~ 100mg kontraktile Proteine [70% MYOSIN, 30% AKTIN] Myosin: ATP-spaltendes Enzym (ATPase) (MG ~ D), besteht aus 2 umeinander verdrillten schweren Peptidketten (heavy chains) und 4 leichten Peptidketten (light chains) Die beiden schweren Ketten bilden am einen Ende 2 globuläre Köpfchen (Subfragment-1)
 (MG ~ D), besteht aus 2 umeinander verdrillten schweren Peptidketten (heavy chains) und 4 leichten Peptidketten (light chains) Die beiden schweren Ketten bilden am einen Ende 2 globuläre Köpfchen (Subfragment-1)")
35
Molekulare Mechanismen der
Muskelkontraktion Myosin: Jedes Köpfchen besitzt eine Bindungsstelle für Aktin, für ATP und für jeweils 2 leichte Ketten Aktin: Globuläres Protein (MG ~ D) wobei die Aktinmonomere (G-Aktin) zu fibrillären Aktinfäden (F-Aktin) polymerisieren MYOFIBRILLEN Aktin und Myosin bilden die dicken und dünnen Filamente der Myofibrillen (innerhalb der Muskelzelle achsenparallel angeordnete kontraktile Organelle) Myofibrillen sind kontraktile Schläuche (~1µm dick)
 wobei die Aktinmonomere (G-Aktin) zu fibrillären Aktinfäden (F-Aktin) polymerisieren. MYOFIBRILLEN Aktin und Myosin bilden die dicken und dünnen Filamente der Myofibrillen (innerhalb der Muskelzelle achsenparallel angeordnete kontraktile Organelle) Myofibrillen sind kontraktile Schläuche (~1µm dick)")
37
Molekulare Mechanismen der
Muskelkontraktion MYOFIBRILLEN Myofibrillen sind durch Trennwände (Z-Scheiben) in zahlreiche Fächer (~ 2.5µm lang) unterteilt (Sarkomere) SARKOMERE erscheinen im Lichtmikroskop (LM) als hell/dunkel gefärbte Bänderung (‚quergestreifte Muskulatur‘); diese entsteht aufgrund einer regel-mäßigen Anordnung der Aktin- und Myosinfilamente. Die Bündel der regelmäßigen ausgerichteten Myosinfilamente (~ 1.6µm lang) erscheinen im LM als dunkle, im polarisierten Licht doppelbrechende (anisotrope) A-Banden
 in zahlreiche Fächer (~ 2.5µm lang) unterteilt (Sarkomere) SARKOMERE erscheinen im Lichtmikroskop (LM) als hell/dunkel gefärbte Bänderung (‚quergestreifte Muskulatur‘); diese entsteht aufgrund einer regel-mäßigen Anordnung der Aktin- und Myosinfilamente. Die Bündel der regelmäßigen ausgerichteten Myosinfilamente (~ 1.6µm lang) erscheinen im LM als dunkle, im polarisierten Licht doppelbrechende (anisotrope) A-Banden.")
38
Schematischer Ausschnitt aus einer Skelettmuskelfaser

39
Molekulare Mechanismen der
Muskelkontraktion Verkürzung der Sarkomere Ein Muskel verkürzt sich durch Übereinandergleiten der Aktin- und Myosinfilamente (Gleitfilamentheorie) Die Verkürzung des Muskels resultiert aus der Verkürzung vieler Sarkomere, die in den Myofibrillen in Serie hintereinandergeschaltet sind. Bei der Verkürzung gleiten die dünnen Aktinfilamente über die dicken Myosinfilamente und schieben sich in Richtung zur Sarkomermitte; bei der Verkürzung werden die Filamente in ihrer Länge NICHT verändert
 Die Verkürzung des Muskels resultiert aus der Verkürzung vieler Sarkomere, die in den Myofibrillen in Serie hintereinandergeschaltet sind. Bei der Verkürzung gleiten die dünnen Aktinfilamente über die dicken Myosinfilamente und schieben sich in Richtung zur Sarkomermitte; bei der Verkürzung werden die Filamente in ihrer Länge NICHT verändert.")
40
A. 2 hintereinander geschaltete Sarkomere unter Ruhebedingung
(erschlaffter Form) B. In verkürzter (kontrahierter) Form
 B. In verkürzter (kontrahierter) Form.")
41
Molekulare Mechanismen der
Muskelkontraktion Funktionsweise der Querbrücken Die Querfortsätze eines Myosinfilaments werden aus den Köpfen von ~ 150 Myosinmolekülen gebildet. Ein jeder Myosinkopf (oder Querfortsatz) kann als Querbrücke ein Myosinfilament mit einem benachbarten Aktinfilament verbinden. Durch eine ‚Kippbewegung‘ der Köpfe werden die Aktinfilamente in Richtung Sarkomermitte gezogen. Aufgrund eines ‚Tauziehprinzips‘ werden in unzähligen in Serie geschalteten Sarkomere die molekularen Bewegungen der Querbrücken in makroskopische Bewegungen umgesetzt
 kann als Querbrücke ein Myosinfilament mit einem benachbarten Aktinfilament verbinden. Durch eine ‚Kippbewegung‘ der Köpfe werden die Aktinfilamente in Richtung Sarkomermitte gezogen. Aufgrund eines ‚Tauziehprinzips‘ werden in unzähligen in Serie geschalteten Sarkomere die molekularen Bewegungen der Querbrücken in makroskopische Bewegungen umgesetzt.")
42
A Modellvorstellung zur Entstehung der Bewegung; A: Myosinfilament mit Querbrücken an benach-barten Aktinfilamenten vor dem Brückenschlag B: Nach dem Ruderschlag der Brücken B Modell für Kraftentstehung in einer Querbrücke vor bzw. nach dem Ruderschlag einer Brücke

43
Molekulare Mechanismen der
Muskelkontraktion Muskelerschlaffung Die Myosinköpfchen lösen sich vom Aktinfaden _> Aktin- u. Myosinfilamente sind dann leicht gegeneinander verschiebbar Als Energiequelle für die Rudertätigkeit der Myosinköpfe wird ATP verwendet; dieses wird durch im Muskel in ADP und Phosphat gespalten (mittels einer Myosin-ATPase) Andere energieliefernde Reaktionen (aerober und anaerober Abbau von KH, Zerfall von KrP) sichern vorwiegend die Neubildung von ATP
 Andere energieliefernde Reaktionen (aerober und anaerober Abbau von KH, Zerfall von KrP) sichern vorwiegend die Neubildung von ATP.")
44
Molekulare Mechanismen der
Muskelkontraktion Wirkungsweise von ATP Nach jedem Ruderschlag wird ein Molekül ATP an das Myosinköpfchen gebunden und liefert damit die Energie für die Trennung von Aktin und Myosin Unmittelbar danach lösen sich die Myosinköpfchen vom Aktin und ATP _> ADP + P (Hydrolyse) Die Enzymprodukte bleiben nach der Spaltung noch gebunden-dies ist die Voraussetzung für das erneute Wiederanfassen der Querbrücke am Aktin und den Querbrückenschlag, bei dem P und dann auch ADP freigesetzt werden.
 Die Enzymprodukte bleiben nach der Spaltung noch gebunden-dies ist die Voraussetzung für das erneute Wiederanfassen der Querbrücke am Aktin und den Querbrückenschlag, bei dem P und dann auch ADP freigesetzt werden.")
45
Molekulare Mechanismen der
Muskelkontraktion Wirkungsweise von ATP Beim Ruderschlag verbiegt sich das Myosinköpfchen und das Aktinfilament wird in Richtung Sarkomermitte gerudert (~6nm) Nach der Abspaltung von ADP kehrt die Querbrücke in ihren Ausgangszustand zurück. Wenn erneut ein ATP-Molekül von der Querbrücke gebunden wird, beginnt mit der Ablösung der Brücke und der erneuten ATP-Hydrolyse ein neuer Quer-brückenzyklus
 Nach der Abspaltung von ADP kehrt die Querbrücke in ihren Ausgangszustand zurück. Wenn erneut ein ATP-Molekül von der Querbrücke gebunden wird, beginnt mit der Ablösung der Brücke und der erneuten ATP-Hydrolyse ein neuer Quer-brückenzyklus.")
46
ATP getriebener Querbrückenzyklus

47
Molekulare Mechanismen der
Muskelkontraktion Querbrückenzyklus Dieser Zyklus wiederholt sich mal/sec. In jedem Arbeitszyklus einer Querbrücke wird einmal ATP gespalten (vermutlich ein Molekül ATP/Querbrücke) Je mehr Querbrücken aktiviert und tätig sind, umso größer ist die ATP-Spaltung pro Zeiteinheit (ATPase Aktivität) und damit die Muskelkraft ATP-Spaltungsrate bzw. Energieumsatzrate und Kraft eines Muskels sind gekoppelt
 Je mehr Querbrücken aktiviert und tätig sind, umso größer ist die ATP-Spaltung pro Zeiteinheit (ATPase Aktivität) und damit die Muskelkraft. ATP-Spaltungsrate bzw. Energieumsatzrate und Kraft eines Muskels sind gekoppelt.")
48
Molekulare Mechanismen der
Muskelkontraktion Karl-Franzens: Totenstarre Sinkt der ATP-Spiegel der Muskelzelle auf 0, so können die Querbrücken nicht mehr gelöst werden; die Querbrücken verbleiben dann im angehefteten Zustand bis zur Autolyse. ATP verhindert od. löst die Starre (Weichmacherwirkung des ATP): Ist ausreichend ATP vorhanden, aber wird die ATP-Spaltung gehemmt (z.B. bei einem Entzug von Ca2+) so können die Querbrücken nicht neu geschlagen werden: Muskelerschlaffung
: Ist ausreichend ATP vorhanden, aber wird die ATP-Spaltung gehemmt (z.B. bei einem Entzug von Ca2+) so können die Querbrücken nicht neu geschlagen werden: Muskelerschlaffung.")
49
Molekulare Mechanismen der
Muskelkontraktion Karl-Franzens: Wirkung von Ca2+ Die Aktivität der Querbrücken wird von der IZ [Ca2+ ] reguliert; liegt IZ [Ca2+ ] < 10-7 mmol/l, so hemmen Regulatorproteine (Troponin, Tropomyosin) den Querbrückenschlag [feste Anbindung der lose gebundenen Querbrücken an das Aktin wird verhindert (Muskel ist relaxiert und kraftlos)]; erhöht sich IZ [Ca2+ ] auf 1-10µM, so werden die lose gebundenen Myosinquerbrücken fest an das Aktin angeheftet
 den Querbrückenschlag [feste Anbindung der lose gebundenen Querbrücken an das Aktin wird verhindert (Muskel ist relaxiert und kraftlos)]; erhöht sich IZ [Ca2+ ] auf 1-10µM, so werden die lose gebundenen Myosinquerbrücken fest an das Aktin angeheftet.")
50
Molekulare Mechanismen der
Muskelkontraktion Karl-Franzens: Wirkung von Ca2+ Die Aktinketten sind in regelmäßigen Abständen mit Troponinmolekülen besetzt, während in den Längsrinnen zwischen den Ketten Fäden aus Tropomyosin laufen. Erst die Bindung von Ca2+ an Troponin (Konformationsänderung des Troponin/Tropomyosinkomplexes) begünstigt und beschleunigt die Bildung stark gebundener Querbrücken
 begünstigt und beschleunigt die Bildung stark gebundener Querbrücken.")
51
Funktionsweisen der Querbrücken (Ca2+)
Wirkung des Ca wichtig. Ca in ruhe bis durch Muskelkontraktion dann angeregt und in dr zelle freigegeben. angeregt. Ca wird an das Troponin gebunden Troponin rutscht weg…Myosinkopf kann andocken.

52
Vorgänge bei der Auslösung einer Muskelkontraktion
Molekulare Mechanismen der Muskelkontraktion Karl-Franzens: Regulation der Muskelkontraktion Vorgänge bei der Auslösung einer Muskelkontraktion Reizung bzw. Erregung der Muskelfaser Aktionspotential (Membranerregung) Elektromechanische Koppelung (Ca Wichtig) Erregungsleitung im T-System (Transversale Kanäle) Kalziumfreisetzung im L-System (Longitudinal) Kalziumwirkung auf die Myofibrillen bzw. Regulatorproteine Kontraktion der Myofibrillen (Querbrückentätigkeit)
 Elektromechanische Koppelung (Ca Wichtig) Erregungsleitung im T-System (Transversale Kanäle) Kalziumfreisetzung im L-System (Longitudinal) Kalziumwirkung auf die Myofibrillen bzw. Regulatorproteine. Kontraktion der Myofibrillen (Querbrückentätigkeit)")
53
Molekulare Mechanismen der
Muskelkontraktion Karl-Franzens: Ca2+ als Botenstoff Kalziumionen spielen eine Schlüsselrolle bei der Übermittlung des Kontraktionssignals von der erregten Zellmembran zu den Myofibrillen also für elekt.mech. Koppelung Die IZ [Ca2+] wird durch das sarkoplasmatische Retikulum (SR) eingestellt (speichert im erschlafften Muskel das Kalzium) dessen Membranen eine ATP-getriebene Kalziumpumpe enthalten; Ca2+ Ionen werden dabei aus dem Myoplasma aktiv in das Innere des longitudinalen Systems transportiert (dadurch wird der myoplasmatische Ca2+ Spiegel auf 10-7 mol/l gesenkt) L-System: Longitudinal Tubussystem (LT) Hier Ca Ionen Pumpe: d.H.: Konzentration im Zellplasma sehr gering > Haftstellen für das Myosinköpchen versperrt.
 eingestellt (speichert im erschlafften Muskel das Kalzium) dessen Membranen eine ATP-getriebene Kalziumpumpe enthalten; Ca2+ Ionen werden dabei aus dem Myoplasma aktiv in das Innere des longitudinalen Systems transportiert (dadurch wird der myoplasmatische Ca2+ Spiegel auf 10-7 mol/l gesenkt) L-System: Longitudinal Tubussystem (LT) Hier Ca Ionen Pumpe: d.H.: Konzentration im Zellplasma sehr gering > Haftstellen für das Myosinköpchen versperrt.")
54
Molekulare Mechanismen der
Muskelkontraktion Karl-Franzens: Elektromechanische Koppelung Das AP breitet sich entlang den Membranen des T-Systems (einstülpung der Zellmembran)in das Innere der Zelle aus, springt auf das L-System über und bewirkt die Freisetzung der Ca2+ Ionen in die Zellflüssigkeit um die Myofibrillen Muskelrelaxation setzt ein, sobald die aktiverenden Ca2+ Ionen wieder in das SR zurückgepumpt werden T-System: Transversale Tubussystem
in das Innere der Zelle aus, springt auf das L-System über und bewirkt die Freisetzung der Ca2+ Ionen in die Zellflüssigkeit um die Myofibrillen. Muskelrelaxation setzt ein, sobald die aktiverenden Ca2+ Ionen wieder in das SR zurückgepumpt werden. T-System: Transversale Tubussystem.")
55
A. : vor dem Reiz: Muskelzelle im Ruhezustand
A.: vor dem Reiz: Muskelzelle im Ruhezustand. Einstülpungen in die Zelle Aussen + innen - => Ruhepotential. Zwischen T-Stülpungen liegen die Terminalzisternen und hohe Calziumkonzentrationen. Dort wo eine Einstülpung liegt liegt eine Trennwand eines Sakromers. Niedrige Calziumkonzentration. B.: 5ms nach Reiz. Aussen – innen + (Umpolarisierung). Diese AP wird über die T-Tubuli getragen…Calzium strömt aus (Durchlässigkeit erhöht) und verteilt sich im Zellplasma C.: nach 20ms, AP ist vorbei und RP ist wieder hergestellt. Aber auströmende Ca hat sich nun verteilt und maximale Konzentration erreicht (100 fache angestiegen)…Myosinköpfchen können nun gebunden werden. Kontraktion steht am Beginn. (nach 200ms hat Kontraktion ein Max. erreicht.) Summation/Superposition von Muskelkontraktionen… Neues AP kann gesetzt werden…einzelne Kontraktionen können summiert werden. WICHTIGES BILDER!!!! Vorraussetzung der erschlaffung: Hineinpumpen der Ca Ionen in T-System bis krit. Konzentration unterschritten ist…
. Diese AP wird über die T-Tubuli getragen…Calzium strömt aus (Durchlässigkeit erhöht) und verteilt sich im Zellplasma. C.: nach 20ms, AP ist vorbei und RP ist wieder hergestellt. Aber auströmende Ca hat sich nun verteilt und maximale Konzentration erreicht (100 fache angestiegen)…Myosinköpfchen können nun gebunden werden. Kontraktion steht am Beginn. (nach 200ms hat Kontraktion ein Max. erreicht.) Summation/Superposition von Muskelkontraktionen… Neues AP kann gesetzt werden…einzelne Kontraktionen können summiert werden. WICHTIGES BILDER!!!! Vorraussetzung der erschlaffung: Hineinpumpen der Ca Ionen in T-System bis krit. Konzentration unterschritten ist…")
56
Molekulare Mechanismen der
Muskelkontraktion Karl-Franzens: Was passiert bei Einzelzuckung und niederfrequenter Summation? Bei der Summation von Einzelzuckungen (ab ~ 10Hz) nehmen zwar die Spannungsmaxima der Kontraktionszyklen und die Kontraktionsrückstände in den aufeinanderfolgenden Zuckungen zu, aber IZ [Ca2+] fällt nach jeder Zuckung fast auf den Ruhewert ab ROT: Intrazelluläre Ca Konzentration BLAU: Mechanogramm Bei 10Hz.: entspannung kommt nicht nach…Summation der Kontraktionen!... Unter 10 Hz. Sinkt es immer wieder zum Ruhewert.
 nehmen zwar die Spannungsmaxima der Kontraktionszyklen und die Kontraktionsrückstände in den aufeinanderfolgenden Zuckungen zu, aber IZ [Ca2+] fällt nach jeder. Zuckung fast. auf den Ruhewert. ab. ROT: Intrazelluläre Ca Konzentration. BLAU: Mechanogramm. Bei 10Hz.: entspannung kommt nicht nach…Summation der Kontraktionen!... Unter 10 Hz. Sinkt es immer wieder zum Ruhewert.")
57
Molekulare Mechanismen der
Muskelkontraktion Karl-Franzens: Was passiert bei Einzelzuckung u. niederfrequenter Summation? Wird die Reizfolge schneller (>20Hz) bleibt der Ca2+ Spiegel zwischen den Reizen erhöht (Kalziumpumpe kann die Ionen nicht vollständig in das L-System des SR zurückpumpen); dann verschmelzen die Einzel- zuckungen (fast) vollständig zum Tetanus (Dauerkontraktion) Tetanische Kontraktion: DAUERKONTRAKTION! Unvollstädengier Tetanus: sieht zacken noch…
 bleibt der Ca2+ Spiegel zwischen den Reizen erhöht (Kalziumpumpe kann die Ionen nicht vollständig in das L-System des SR zurückpumpen); dann verschmelzen die Einzel- zuckungen (fast) vollständig. zum Tetanus. (Dauerkontraktion) Tetanische Kontraktion: DAUERKONTRAKTION! Unvollstädengier Tetanus: sieht zacken noch…")
58
Molekulare Mechanismen der
Muskelkontraktion Tetanusverschmelzungsfrequenz Einzelzuckungen verschmelzen zum vollständigen Tetanus wenn das Reizintervall kleiner als ~1/3 der für die Einzelzuckung benötigten Zeit ist; die Tetanusverschmelzungsfrequenz (TVF) ist umso niedriger, je länger die Einzelzuckung dauert, und sie ist temperatur abhängig. Wird die Stimulationsrate über die TVF hinaus erhöht, nimmt die [Ca2+] zu und damit auch die Muskelkraft TVF: Wenn man weiter erhöht…kann nur noch Konzentrationskraft erhöht werde bis zur ermüdung
 ist umso niedriger, je länger die Einzelzuckung dauert, und sie ist temperatur abhängig. Wird die Stimulationsrate über die TVF hinaus erhöht, nimmt die [Ca2+] zu und damit auch die Muskelkraft. TVF: Wenn man weiter erhöht…kann nur noch Konzentrationskraft erhöht werde bis zur ermüdung.")
59
Molekulare Mechanismen der
Muskelkontraktion Bei einem zur Ermüdung führenden Tetanus nimmt die [Ca2+] und die Muskelkraft wieder ab, der minimale zeitliche Abstand zwischen aufeinanderfolgenden effektiven Reizen im Tetanus kann nicht kleiner sein, als die Refraktärzeit (~ Dauer eines AP) - wird eine Kontraktion ohne AP‘s ausgelöst (z.B. Koffein): Kontraktur Kontraktur (Gelenksversteifung…nicht über AP oder motorische Nervensystem ausgelöste Muskelkontraktionen…wenn zu viel Ca ausgeschüttet wird durch Koffein) ist nicht das selbe wie Tetanus Krampf wird auch über das Nervensystem ausgelöst
 - wird eine Kontraktion ohne AP‘s ausgelöst (z.B. Koffein): Kontraktur. Kontraktur (Gelenksversteifung…nicht über AP oder motorische Nervensystem ausgelöste Muskelkontraktionen…wenn zu viel Ca ausgeschüttet wird durch Koffein) ist nicht das selbe wie Tetanus. Krampf wird auch über das Nervensystem ausgelöst.")
60
Molekulare Mechanismen der Regulation der Muskelkraft
Muskelkontraktion Regulation der Muskelkraft Motorische Einheit (ME) Wird gebildet aus einem Motoneuron und dem von ihm innervierten Muskelfasern. ME‘s sind unterschiedlich groß; dort wo die Feinmotorik wichtig ist (z.B. Augenmuskulatur, Finger-u.Handmuskulatur) versorgt ein Motoneuron nur wenige MF; dort, wo die Feinmotorik keine wichtige Rolle spielt aber unter Umständen große Kraftleistungen vollbracht werden, innerviert ein MN sehr viele MF: ‚Innervationsverhältnis‘ ÜBERBLICK: Zunehmende Zahl von motor. Einheiten werden aktiviert (je mehr gleichzeitig rekrotiert werden desto größer ist die entwickelte Kraft…untrainierte Muskel kann nur 60% dieser ME verwenden…wird aber schnell aufgebaut (Tage)) Innervationsverhätnis: Feinmotorik (ME klein) Bsp. Augen…eine ME mit 10 fasern…sehr genau!, statische Kraft (ME groß, ein Motorneuron hat viele Fasern. Rückenmuskulatur )
 Wird gebildet aus einem Motoneuron und dem von ihm innervierten Muskelfasern. ME‘s sind unterschiedlich groß; dort wo die Feinmotorik wichtig ist (z.B. Augenmuskulatur, Finger-u.Handmuskulatur) versorgt ein Motoneuron nur wenige MF; dort, wo die Feinmotorik keine wichtige Rolle spielt aber unter Umständen große Kraftleistungen vollbracht werden, innerviert ein MN sehr viele MF: ‚Innervationsverhältnis‘ ÜBERBLICK: Zunehmende Zahl von motor. Einheiten werden aktiviert (je mehr gleichzeitig rekrotiert werden desto größer ist die entwickelte Kraft…untrainierte Muskel kann nur 60% dieser ME verwenden…wird aber schnell aufgebaut (Tage)) Innervationsverhätnis: Feinmotorik (ME klein) Bsp. Augen…eine ME mit 10 fasern…sehr genau!, statische Kraft (ME groß, ein Motorneuron hat viele Fasern. Rückenmuskulatur )")
61
Molekulare Mechanismen der Regulation der Muskelkraft
Muskelkontraktion Regulation der Muskelkraft Motorische Einheit (ME) Die Kraft varriert bei einer Einzelzuckung nur gering (infolge des Alles oder Nichts Prinzips) weil in der Einheit entweder alle Fasern entweder gereizt od. erschlafft sind- Wie kann nun die Kraft beeinflußt werden? Rekrutierung ME‘s 0/1 gesetzt: Reicht das ATP aus den Schwellwert zu überschreiten oder nicht? Mehr ME Summation Steigerung der Entladungsfrequenz der -Motoneuronen
 Die Kraft varriert bei einer Einzelzuckung nur gering (infolge des Alles oder Nichts Prinzips) weil in der Einheit entweder alle Fasern entweder gereizt od. erschlafft sind- Wie kann nun die Kraft beeinflußt werden Rekrutierung ME‘s. 0/1 gesetzt: Reicht das ATP aus den Schwellwert zu überschreiten oder nicht Mehr ME. Summation. Steigerung der Entladungsfrequenz der -Motoneuronen.")
62
Molekulare Mechanismen der Regulation der Muskelkraft
Muskelkontraktion Regulation der Muskelkraft Rekrutierung ME‘s Die Muskelkraft u. die Kontraktionsgeschwindigkeit können durch die Aktivierung (Rekrutierung) von immer mehr ME‘s gesteigert werden - die Feinregulierung der Kraft ist dabei umso besser abstufbar, je geringer die Größe (und damit die Kraft) einer ME ist. Bei geringer Willküranspannung produzieren nur sehr wenige ME‘s AP, bei starker Anstrengung feuern sehr viele ME‘s Je geringer die Größe einer ME bzw umso kleiner das Innervationsverhältnis.!
 von immer mehr ME‘s gesteigert werden - die Feinregulierung der Kraft ist dabei umso besser abstufbar, je geringer die Größe (und damit die Kraft) einer ME ist. Bei geringer Willküranspannung produzieren nur sehr wenige ME‘s AP, bei starker Anstrengung feuern sehr viele ME‘s. Je geringer die Größe einer ME bzw umso kleiner das Innervationsverhältnis.!")
63
Molekulare Mechanismen der Regulation der Muskelkraft
Muskelkontraktion Regulation der Muskelkraft Steigerung der Entladungsfrequenz der -MN Die Änderung der Stimulierungsfrequenz beeinflußt die Kraft; aufgrund der Überlagerung u. des Summationseffekts ist die Kraft im vollständigen Tetanus (bei einer hohen Impulsrate der -MN) mindestens doppelt so groß wie in einem unvollständig verschmolzenen Tetanus mit niedriger SF. Durch Steigerung der Impulsrate der MN entwickelt sich ein verschmolzener, glatter Tetanus wodurch sich die Kontraktionskraft auf den 2- bis 10-fachen Wert erhöht Praktisch für alle Bew. gibt es immer tetanische Kontraktionen Spezialfall: Kniesehnenreflexe (Patellasehne!)
 mindestens doppelt so groß wie in einem unvollständig verschmolzenen Tetanus mit niedriger SF. Durch Steigerung der Impulsrate der MN entwickelt sich ein verschmolzener, glatter Tetanus wodurch sich die Kontraktionskraft auf den 2- bis 10-fachen Wert erhöht. Praktisch für alle Bew. gibt es immer tetanische Kontraktionen. Spezialfall: Kniesehnenreflexe (Patellasehne!)")
64
Molekulare Mechanismen der Regulation der Muskelkraft
Muskelkontraktion Regulation der Muskelkraft „Sonderfall“ Synchronisation In Folge der Anpassung an spezifisches Krafttraining (v.a. Maximalkrafttraining und Schnellkraftraining) werden im Zuge einer Kraftanstrengung bzw. Belastung verschieden große ME gleichzeitig aktiviert – damit wird in kurzer Zeit viel Kraft entwickelt (hängt aber wiederum von der Impulsfrequenz ab) Intramuskuläre Koordinationsverbesserung
 werden im Zuge einer Kraftanstrengung bzw. Belastung verschieden große ME gleichzeitig aktiviert – damit wird in kurzer Zeit viel Kraft entwickelt (hängt aber wiederum von der Impulsfrequenz ab) Intramuskuläre Koordinationsverbesserung.")
65
EMG Elektromyogram Elektromyography
AP‘s infolge De- und Repolarisationsvorgänge

66
EMG Elektromyogram Elektromyography
Dipol v Muskel als Dipol Dipol wandert in der Muskelfaser = Fortleitung Muskelfaseroberfläche

67
EMG Elektromyogram Elektromyography
d Elektrode + - d‘ P D Innen Negativ aussen Positiv Muskelfaseroberfläche: Dipolvektor an der Front der Errengungswelle (Ausschlag ins Negative) Dipolvektor am Ende der Erregungswelle (Ausschlag ins Positive) Extrazelluläre Ableitung: Elektromyogramm (EMG) 1 2 3 4 5 Muskelfaseroberfläche
 Dipolvektor am Ende der Erregungswelle (Ausschlag ins Positive) Extrazelluläre Ableitung: Elektromyogramm (EMG) Muskelfaseroberfläche.")
68
EMG Elektromyogram Elektromyography
Signalaufnahme Oberflächen- Dünndrahtelektroden Oberflächen Elektroden: Erfassung von Potentialänderungen auf der Hautoberfläche: extra-zelluläre Summenpotentiale eines Musekls oder einer Muskelgruppe. Dünndraht Elektroden: Erfassung von extrazellulären Potentialschwankungen einer motorischen Einheit erfassen

69
EMG Elektromyogram Elektromyography

70
EMG Elektromyogram Elektromyography

71
EMG Elektromyogram Elektromyography
Aktivität eines Muskels

72
EMG Elektromyogram Elektromyography
Kniestreckung: Max = Streckung, Min = Beugung

73
EMG Elektromyogram Elektromyography

74
Molekulare Mechanismen der Regulation der Muskelkraft
Muskelkontraktion Regulation der Muskelkraft Muskelfasertypen Die Farbe eines Muskels ist weitgehend durch den Gehalt an Myoglobin (Protein, dass der O2 Aufnahme und Bindung in der Muskelzelle dient) Generell lassen sich 2 Typen von MF unterscheiden: Weiße MF (myoglobinarm) Rote MF (myoglobinreich) Myoglobin: Muskelfarbstoff, soll Sauerstoff aufnehmen und speicher und bei Bedarf wieder abgeben. Myoglobin ist aber in der Muskelzelle inkludiert und liegt nicht frei wie das Hämoglobin (roten Blutkörperchen…diese Transportiert auch Sauerstoff) 2 Typen: Weiße MF, Rote MF (können Sauerstoff besser speichern und Verwerten!) Oxidativ: aerob Glykolytisch: anaerob Die Fasern dieser Muskeln sind strukturell, funktionell und biochemisch unterschiedlich (v.a. der Gehalt an Enzymen des oxidativen bzw glykolitischen Stoffwechsels, und die Aktivität der Myosin-ATPase)
 Generell lassen sich 2 Typen von MF unterscheiden: Weiße MF (myoglobinarm) Rote MF (myoglobinreich) Myoglobin: Muskelfarbstoff, soll Sauerstoff aufnehmen und speicher und bei Bedarf wieder abgeben. Myoglobin ist aber in der Muskelzelle inkludiert und liegt nicht frei wie das Hämoglobin (roten Blutkörperchen…diese Transportiert auch Sauerstoff) 2 Typen: Weiße MF, Rote MF (können Sauerstoff besser speichern und Verwerten!) Oxidativ: aerob. Glykolytisch: anaerob. Die Fasern dieser Muskeln sind strukturell, funktionell und biochemisch unterschiedlich (v.a. der Gehalt an Enzymen des oxidativen bzw glykolitischen Stoffwechsels, und die Aktivität der Myosin-ATPase)")
75
Molekulare Mechanismen der Regulation der Muskelkraft
Muskelkontraktion Karl-Franzens: Regulation der Muskelkraft Rote Muskulatur findet sich vorwiegend in der Rumpf/Stützmuskulatur (Typ-1 Muskulatur, langsame MF, niedrige Myosin-ATPase-Aktivität) Vorteil: ermüdungsresistent Nachteil: keine sehr hohe und schnelle Kraftentwicklung Farbe rot Kontraktionsform langsame Zuckung Ermüdbarkeit gering Stoffwechsel oxidativ (aerob) Myosin-ATPase niedrig LDH-Aktivtät niedrig Roten Muskelfaser: Ausdauermuskeln Lactate-Hydrogenase : Produziert Lactat und baut auch ab…langsame Kontraktion kein Lactat
 Vorteil: ermüdungsresistent. Nachteil: keine sehr hohe und schnelle Kraftentwicklung. Farbe rot. Kontraktionsform langsame Zuckung. Ermüdbarkeit gering. Stoffwechsel oxidativ (aerob) Myosin-ATPase niedrig. LDH-Aktivtät niedrig. Roten Muskelfaser: Ausdauermuskeln. Lactate-Hydrogenase : Produziert Lactat und baut auch ab…langsame Kontraktion kein Lactat.")
76
Molekulare Mechanismen der Regulation der Muskelkraft
Muskelkontraktion Regulation der Muskelkraft Weiße Muskulatur findet sich dort, wo ballistische Bewegungen vermittelt werden (z.B. Körperfern) (Typ-2a u. Typ 2b MF, schnelle u. kraftvolle MF, hohe Myosin-ATPase-Aktivität) Vorteil: hohe und schnelle Kraftbildungsrate Nachteil: rasche Ermüdung IIa IIb Farbe rosa weiß Kontraktionsform schnell schnellste Ermüdbarkeit mittel rasch Stoffwechsel oxid/glykol. glykolytisch Myosin-ATPase hoch hoch LDH-Aktivtät mittel hoch
 (Typ-2a u. Typ 2b MF, schnelle u. kraftvolle MF, hohe Myosin-ATPase-Aktivität) Vorteil: hohe und schnelle Kraftbildungsrate. Nachteil: rasche Ermüdung. IIa IIb. Farbe rosa weiß. Kontraktionsform schnell schnellste. Ermüdbarkeit mittel rasch. Stoffwechsel oxid/glykol. glykolytisch. Myosin-ATPase hoch hoch. LDH-Aktivtät mittel hoch.")
77
Molekulare Mechanismen der Regulation der Muskelkraft
Muskelkontraktion Regulation der Muskelkraft Bedeutung V.a. in Sportarten mit hohen Kraft- u. Schnelligkeitsanteilen ist die Dominanz der IIb MF wichtig - umgekehrt werden Sportarten, die durch einen hohen Anteil aerober Energiebereitstellung gekennzeichnet sind, eher von der Dominanz des Typ I profitieren MF-Verteilung ist in hohem Maße genetisch determiniert und kann durch ein entsprechendes Training kaum nur wenig verändert werden

78
Molekulare Mechanismen der Regulation der Muskelkraft
Muskelkontraktion Regulation der Muskelkraft Umwandlungsmöglichkeiten sind nur mittels der Kreuzinnervation möglich (eine langsame MF bzw. ein langsamer Muskel wird von einem großen, schnellen MN mit entsprechend hoher Frequenz innerviert – dabei lassen sich morphologische Veränderungen in Richtung schneller MF erzielen-bei Wegfall der hohen Impulsrate erfolgt aber wieder die Reanpassung) Kreuzinnervation: Tierexperiment….kann theoretisch die Fasern umändern in beide Richtungen! Sprinter => Marathon Läufer leichter als umgekehrt Aber: es ist leichter, eine schnelle MF in eine langsame zu transformieren ……..
 Kreuzinnervation: Tierexperiment….kann theoretisch die Fasern umändern in beide Richtungen! Sprinter => Marathon Läufer leichter als umgekehrt. Aber: es ist leichter, eine schnelle MF in eine langsame zu transformieren ……..")
79
Molekulare Mechanismen der Regulation der Muskelkraft
Muskelkontraktion Regulation der Muskelkraft Muskelmechanik Kontraktionsformen der Muskulatur Isometrische Kontraktion (gleichbleibende Länge) Konzentrische Kontraktion (Verkürzung) Exzentrische Kontraktion (Dehnung im Zuge einer Anspannung) (isotonische-, auxotonische Kontraktion) gleichbleibende Spannung bzw ~ gleichbleibende Spannung unter Verkürzung Muskelfasern: 1 <> 2C <> 2A <> 2B (1 auf 2B geht ned!) Langsam (S,s) zu schnell (F,f) S, S+Fa, Fa, Fb Vorkommen langsamer (S,s) und schneller (F,f) Myosinmolküle mit den entspre3chenden schweren (HC) und leichten (LH)… Tabelle: Kontraktionszeit/… Bild: zentralnervensystem: Stützmotorik (ExtraPyramdalMotorisches System, EPMS; 10 Hz), Zielmotorik - Willkürlich (PMS, 40 Hz) 2 Arten von Muskelkontraktionen: - Isometrisch: gleichermaßen: statische (halten,stehen) Kontraktion; Spannungszunahme bei gleichbleibender Länge - Isotonisch: verkürzung bei gleicher Spannung (schlecht für schwere Gewichte) - Auxotonisch: gleichzeitige Änderung von Spannung und Länge Kontraktionsformen: - statisch Kontraktion - dynamische Kontraktio n - positiv (konzentrische) dynamisch: isotonische Verkürzung - negativ (exzentrisch)dynamisch: dehnung des Muskels im Zuge einer Anspannung (Geischt zu schwer…kine Verkürzung…wird gedehnt während dem anspannen)
 Konzentrische Kontraktion (Verkürzung) Exzentrische Kontraktion (Dehnung im Zuge einer Anspannung) (isotonische-, auxotonische Kontraktion) gleichbleibende Spannung bzw ~ gleichbleibende Spannung unter Verkürzung. Muskelfasern: 1 <> 2C <> 2A <> 2B (1 auf 2B geht ned!) Langsam (S,s) zu schnell (F,f) S, S+Fa, Fa, Fb. Vorkommen langsamer (S,s) und schneller (F,f) Myosinmolküle mit den entspre3chenden schweren (HC) und leichten (LH)… Tabelle: Kontraktionszeit/… Bild: zentralnervensystem: Stützmotorik (ExtraPyramdalMotorisches System, EPMS; 10 Hz), Zielmotorik - Willkürlich (PMS, 40 Hz) Arten von Muskelkontraktionen: - Isometrisch: gleichermaßen: statische (halten,stehen) Kontraktion; Spannungszunahme bei gleichbleibender Länge. - Isotonisch: verkürzung bei gleicher Spannung (schlecht für schwere Gewichte) - Auxotonisch: gleichzeitige Änderung von Spannung und Länge Kontraktionsformen: - statisch Kontraktion. - dynamische Kontraktio n. - positiv (konzentrische) dynamisch: isotonische Verkürzung. - negativ (exzentrisch)dynamisch: dehnung des Muskels im Zuge einer Anspannung (Geischt zu schwer…kine Verkürzung…wird gedehnt während dem anspannen)")
80
Molekulare Mechanismen der
Muskelkontraktion Muskelmechanik Ruhedehnungskurve Die Muskelkraft hängt von der aktuellen Länge des Muskels ab, der ruhende Muskel verhält sich elastisch jedoch nimmt bei der Dehnung die Spannung nicht linear zu (im Ggs. zu einer Feder) – das Elastizitätsmodul des ruhenden Muskels nimmt somit mit der Dehnung zu Die Elastizität kommt hptsl. durch dehnbare Strukturen zustande, die zu den kontraktilen Elementen parallel geschaltet sind (parallelgeschaltete Elemente; Sarkolemm, bindegewebige Strukturen zwischen den Fasern und Titin) : W.H.: fuskelfasertypen und deren umwandlung in verschiedene Typen. Feststellung der Myosinmoleküle durch Elektrostimulation Typ I: 10 AP/sec, Typ II: 40AP/sec Umwandlung vie Kreuzinversion…langsamen nerven abgetrennt und schnellen angeschlossen…geht auch vice versa. Geht in beide Richtungen (vollständig). Typ I > Typ II: geht schlecht Typ II > Typ I: geht besser Typ I (stützmotorik, unbewußt) , TypII (zielmotorik) Krezinversion << >> Elektrostimulation << > Ausdauertraining: < Krafttraining: > Intervalltraining: > Ruhedehnungskurve: Passive elastische Eigenschaften Muskelkraft hängt von der Länge ab Spannung nimmt nicht wie bei der Stahlfeder linear sondern exponentiell Elastizitätsmodul (Verhältnis von Spannung und Dehnung) Parallelelastische elemente: tertiären filamente bzw. Proteine W.H.: 3 Arten von Filamenten: Myosin Tropomin (kontraktion…), aktin usw. Tertiäre elastische filament: Titinfilamente (um Myosin 6:1) Nebulinfilamente: um Aktin Longitudinale und transversale intermediäre Filamente inkl. Desmin Kurze Filamente zur Verbindung von Muskel und Sehnenfaser (integrin: Dystrophin. Talin. Vinculin…) An der passiv elastischen eigenschaften des Muskels
 – das Elastizitätsmodul des ruhenden Muskels nimmt somit mit der Dehnung zu. Die Elastizität kommt hptsl. durch dehnbare Strukturen zustande, die zu den kontraktilen Elementen parallel geschaltet sind (parallelgeschaltete Elemente; Sarkolemm, bindegewebige Strukturen zwischen den Fasern und Titin) : W.H.: fuskelfasertypen und deren umwandlung in verschiedene Typen. Feststellung der Myosinmoleküle durch Elektrostimulation. Typ I: 10 AP/sec, Typ II: 40AP/sec. Umwandlung vie Kreuzinversion…langsamen nerven abgetrennt und schnellen angeschlossen…geht auch vice versa. Geht in beide Richtungen (vollständig). Typ I > Typ II: geht schlecht. Typ II > Typ I: geht besser. Typ I (stützmotorik, unbewußt) , TypII (zielmotorik) Krezinversion << >> Elektrostimulation << > Ausdauertraining: < Krafttraining: > Intervalltraining: > Ruhedehnungskurve: Passive elastische Eigenschaften. Muskelkraft hängt von der Länge ab. Spannung nimmt nicht wie bei der Stahlfeder linear sondern exponentiell. Elastizitätsmodul (Verhältnis von Spannung und Dehnung) Parallelelastische elemente: tertiären filamente bzw. Proteine. W.H.: 3 Arten von Filamenten: Myosin Tropomin (kontraktion…), aktin usw Tertiäre elastische filament: Titinfilamente (um Myosin 6:1) Nebulinfilamente: um Aktin. Longitudinale und transversale intermediäre Filamente inkl. Desmin. Kurze Filamente zur Verbindung von Muskel und Sehnenfaser (integrin: Dystrophin. Talin. Vinculin…) An der passiv elastischen eigenschaften des Muskels.")
81
Molekulare Mechanismen der
Muskelkontraktion Muskelmechanik Isometrische Maxima Die Vordehnung bestimmt nicht nur die Größe der passiv elastischen Anspannung sondern auch das Ausmaß der Kraft, welches der Muskel bei der jeweiligen Länge entwickeln kann, wenn er aktiviert ist

82
Serienelastisches Element und Parallel Elastisches Element
So erhalte ich die Ruhelängediagramme Dann mittels elektrischen Impulse erzeuge ich isometrische Kontraktionen!!!

83
Ruhelänge: L0 (passive Ruhespannung, anstieg erst jenseits der Ruhelänge des Muskel)
Erst nach bestimmten Länge steigt die Spannung Gelb: Ruhedehnung/Spannungskurve: Ausgangslage für jede aktive Kontraktion A>B: isometrisch: Länge bleibt gleich, Spannung nimmt zu In der Waagrechten: isotonisch: verkürzung bei gleichbleibender Spannung Auxotonisch wäre A zu B: Verkürzung + Spannung Speziallfall von auxotonisch: Verkürzung/Spannung hintereinander ned gleichzeitig. Unterstützungskontraktion: Phase 1: isometerische Anfangsphase und Phase 2: isotonische Endphase Sonst würde das Gewicht den Muskel dehnen! 2ter spezialfall: Anschlagkontraktion: Isotonischer anfangsphase (Hubphase) und Isometrische Endphase....hebt etwas bis zum anschlag…Muskelkönnte weiter braucht aber ned In verschiedenen Muskellngenstadien werden isometrische Kontraktionen hervorgerufen Bei punkt L0 Liegt die Maximale Spannung (Optimallänge) a: Ruhedehnungs Ruhespannungskurve b: Kurve der Isometrischen Maximas AB CD 0E: unterschiedliche maximale isometrische Kontraktionen Ziehe von der Gesamtspannung die passive Spannung ab: Differenzkurve = aktive Spannung Sieht von anfang bis ende wie eine Glockenkurve aus L0: ist die Ruheläng oder funktionelle Länge eines Muskels. In deisem Längenzustand entwickelt ein Muskel seine größte aktive isometrische Spannung…
 und Isometrische Endphase....hebt etwas bis zum anschlag…Muskelkönnte weiter braucht aber ned. In verschiedenen Muskellngenstadien werden isometrische Kontraktionen hervorgerufen. Bei punkt L0 Liegt die Maximale Spannung (Optimallänge) a: Ruhedehnungs Ruhespannungskurve. b: Kurve der Isometrischen Maximas. AB CD 0E: unterschiedliche maximale isometrische Kontraktionen. Ziehe von der Gesamtspannung die passive Spannung ab: Differenzkurve = aktive Spannung. Sieht von anfang bis ende wie eine Glockenkurve aus. L0: ist die Ruheläng oder funktionelle Länge eines Muskels. In deisem Längenzustand entwickelt ein Muskel seine größte aktive isometrische Spannung…")
84
Molekulare Mechanismen der
Muskelkontraktion Muskelmechanik Sarkomerlängen-Kraft Beziehung Man findet ein charakteristisches Optimum bei einer Sarkomerlänge von 2-2.2µm (entspricht ~ der Muskelruhelänge) Bei kleineren Muskellängen ist die Kraft geringer, weil sich die Aktin u. Myosinfilamente gegenseitig behindern u. weil die elektromechan. Koppelung beeinträchtigt ist (die meisten Muskeln können sich nur auf ~ 50-70% ihrer Ruhelänge verkürzen) Größtes Kraftmaximum: bei ym
 Bei kleineren Muskellängen ist die Kraft geringer, weil sich die Aktin u. Myosinfilamente gegenseitig behindern u. weil die elektromechan. Koppelung beeinträchtigt ist (die meisten Muskeln können sich nur auf ~ 50-70% ihrer Ruhelänge verkürzen) Größtes Kraftmaximum: bei ym.")
85
Sarkomerlängen-Kraft Beziehung

86
Molekulare Mechanismen der
Muskelkontraktion Muskelmechanik Sarkomerlängen-Kraft Beziehung Werden MF über ihre Ruhelänge hinaus gedehnt, so fällt die Kontraktionskraft ab, weil die Aktinfilamente aus der Anordnung der Myosinfilamente herausrutschen [eine immer geringer werdende Zahl an Myosinköpfchen bildet mit Aktin eine immer geringer werdende Zahl an Querbrücken] Ebenso ist der durch die Elastizität der Querbrücken bedingte dynamische Dehnungswiderstand („Stiffness“) reduziert – wenn die Sarkomere über ~ 3.6µm hinaus gedehnt werden, können die Myofibrillen keine aktive Kraft mehr entwickeln Weil Myosinfilamente aus den Aktinfilamenten herausgezogen werden
 reduziert – wenn die Sarkomere über ~ 3.6µm hinaus gedehnt werden, können die Myofibrillen keine aktive Kraft mehr entwickeln. Weil Myosinfilamente aus den Aktinfilamenten herausgezogen werden.")
87
Molekulare Mechanismen der
Muskelkontraktion Muskelmechanik Verkürzungsgeschwindigkeit Bei einer tetanischen Aktivierung eines Muskels hängt die Geschwindigkeit einer isoton. Kontraktion (vk) von der Belastung ab – vk nimmt mit steigender Belastung ab („Hill‘sche Kraft-Geschwindigkeitsrelation“ ) Unbelastet verkürzt sich ein Muskel mit seiner maximalen vk wobei die max. vk eines Sarkomers der max. v des Übereinandergeleitens der Aktin- u. Myosinfilamente entspricht (je schneller die Myosinquerbrücken ATP spalten, und mit Aktin wechselwirken, desto höher ist die v des Gleitprozesses)
 von der Belastung ab – vk nimmt mit steigender Belastung ab („Hill‘sche Kraft-Geschwindigkeitsrelation ) Unbelastet verkürzt sich ein Muskel mit seiner maximalen vk wobei die max. vk eines Sarkomers der max. v des Übereinandergeleitens der Aktin- u. Myosinfilamente entspricht (je schneller die Myosinquerbrücken ATP spalten, und mit Aktin wechselwirken, desto höher ist die v des Gleitprozesses)")
88
Molekulare Mechanismen der
Muskelkontraktion Muskelmechanik Bei gleicher vk einzelner Sarkomere kontrahieren lange Muskeln schneller, als kurze (die Verkürzungen der hintereinander geschalteten Sarkomere addieren sich) Last-Geschwindigkeitsbeziehung Die vk nimmt mit zunehmender Last hyperbolisch ab – die vk beträgt etwa 20% der max. möglichen vk (bei lastfreier Verkürzung), wenn die relative Muskelbelastung ~ 50% der max. möglichen unter isometrischen Bedingungen erreichten Kraft beträgt
 Last-Geschwindigkeitsbeziehung. Die vk nimmt mit zunehmender Last hyperbolisch ab – die vk beträgt etwa 20% der max. möglichen vk (bei lastfreier Verkürzung), wenn die relative Muskelbelastung ~ 50% der max. möglichen unter isometrischen Bedingungen erreichten Kraft beträgt.")
90
Molekulare Mechanismen der
Muskelkontraktion Muskelmechanik Last-Geschwindigkeitsbeziehung Ist die Belastung gleich groß wie die isometrisch mögliche Kraft, so verkürzt sich der Muskel nicht; steigt die Belastung über diesen Wert, so wird er gedehnt (exzentrische Belastung hohe Spannungsspitzen, die zu Läsionen im Bereich der z-Scheiben führen können: Muskelkater) Aber: im Zuge eines exzentrisch-orientierten Krafttrg. kommt es zu hohen Kraftzuwachsraten innerhalb kurzer Trgs.perioden (Vermutung: dem ZNS fehlen „Bewegungsprogramme“ für die Exzentrik)
 Aber: im Zuge eines exzentrisch-orientierten Krafttrg. kommt es zu hohen Kraftzuwachsraten innerhalb kurzer Trgs.perioden (Vermutung: dem ZNS fehlen „Bewegungsprogramme für die Exzentrik)")
91
Molekulare Mechanismen der
Muskelkontraktion Muskelmechanik Kraft-Geschwindigkeitsbeziehung Während einer schnellen Verkürzung entwickelt ein Muskel weniger kontraktile Kräfte als bei langsamer Verkürzung od. Dehnung Ausführung von sehr schnelle, leichten Bewegungen nur bei geringer Kraftaufwendung (bei entspannter Muskulatur) Die größte Muskelkraft wird bei langsamen Bewegungen erreicht (schwere Gegenstände können nur langsam gehoben werden)
 Die größte Muskelkraft wird bei langsamen Bewegungen erreicht (schwere Gegenstände können nur langsam gehoben werden)")
92
Molekulare Mechanismen der
Muskelkontraktion Muskelmechanik Kraft-Geschwindigkeitsbeziehung Wenn ein Muskel mit allen zur Verfügung stehenden Fasern eine Last bewältigt, so ist die relative Belastung der einzelnen tätigen MF kleiner und daher deren vk größer als wenn nur ein Teil der MF tätig ist Die Kontraktionsgeschwindigkeit (und Muskel-leistung) kann durch Rekrutierung von ME‘s erhöht werden (und vice versa) Muskelleistung Produkt aus Muskelkraft und Verkürzungs-geschwindigkeit
 kann durch Rekrutierung von ME‘s erhöht werden (und vice versa) Muskelleistung. Produkt aus Muskelkraft und Verkürzungs-geschwindigkeit.")
93
Exkurs-Krafttraining
Anpassungserscheinungen im Krafttraining 1. Die vorrangigen Mechanismen in einer Verbesserung der muskulären Kraft sind neuronal bedingt intramuskulär/intermuskulär 2. Infolge eines längerfristigen KT ist in zunehmendem Maße eine Muskelhypertrophie die Grundlage für den Kraftzuwachs [Zumindest experimentell lässt sich eine Muskelhyperplasie nachweisen – ob diese auch als Folge eines KT beim Menschen auftritt, ist nicht eindeutig geklärt]

94
Die neuronalen Anpassungen hängen ab von:
Exkurs-Krafttraining Anpassungserscheinungen im Krafttraining Die neuronalen Anpassungen hängen ab von: a. Intensität der Belastung (ab ~ 90% der individuellen Maximalkraft) b. Ausführung der Bewegung (Bewgs-geschwindigkeit) c. Anzahl der Wh innerhalb einer Serie und Anzahl der Serien insgesamt d. Belastungspause
 b. Ausführung der Bewegung (Bewgs-geschwindigkeit) c. Anzahl der Wh innerhalb einer Serie und Anzahl der Serien insgesamt. d. Belastungspause.")
95
Eine Muskelhypertrophie hängt ab von:
Exkurs-Krafttraining Anpassungserscheinungen im Krafttraining Eine Muskelhypertrophie hängt ab von: a. Intensität der Belastung (~ 60% - 85% der individuellen Maximalkraft; also Krafteinsatz gegen submaximale Lasten) Anzahl der Wh innerhalb einer Serie (Wh > 5 u. < 15 Wh) und Anzahl der Serien insgesamt Belastungspause Sicherstellung einer Ausbelastung der trainierten Muskulatur innerhalb einer Serie (Einwirkzeit)
 Anzahl der Wh innerhalb einer Serie. (Wh > 5 u. < 15 Wh) und Anzahl der Serien insgesamt. Belastungspause. Sicherstellung einer Ausbelastung der trainierten Muskulatur innerhalb einer Serie (Einwirkzeit)")
96
Exkurs-Krafttraining
Allgemeine Auswirkungen eines Krafttrainings Verbesserung der Stoffwechselsituation (v.a. Steigerung der Glukose- u. Insulinsensitivität durch Änderung der Rezeptorenschwelle, Zunahme der Anzahl von Rezeptoren bzw Beeinflußung der intrazellulären Signalkaskade) und der Sensitivität gegenüber anderen endokrinen Faktoren (Wachstumsfaktoren, wie IGF‘s, Bindungsproteine, etc.)
 und der Sensitivität gegenüber anderen endokrinen Faktoren (Wachstumsfaktoren, wie IGF‘s, Bindungsproteine, etc.)")
97
Exkurs-Krafttraining
Allgemeine Auswirkungen eines Krafttrainings 2. Zunahme der Muskelmasse (LBM, lean body mass) führt zu einer Erhöhung des Grundumsatzes (Adipositas‘vorbeugung‘) 3. KT im Alter wirkt dem Muskelabbau (Sarkopenie) entgegen und wirkt auch im Sinne einer Osteoporoseprophylaxe (verbesserter Knochenstoffwechsel) 4. KT führt zu einer Verbesserung v.a. der Stützfunktion entsprechender Muskelgruppen und kann damit Haltungsproblemen bzw Bewegungsmangelkrankheiten vorbeugen
 führt zu einer Erhöhung des Grundumsatzes (Adipositas‘vorbeugung‘) 3. KT im Alter wirkt dem Muskelabbau (Sarkopenie) entgegen und wirkt auch im Sinne einer Osteoporoseprophylaxe (verbesserter Knochenstoffwechsel) 4. KT führt zu einer Verbesserung v.a. der Stützfunktion entsprechender Muskelgruppen und kann damit Haltungsproblemen bzw Bewegungsmangelkrankheiten vorbeugen.")
98
Nach M.J. Rennie: J. of Physiology 2001

99
Michael J. Rennie J. of Physiology 2001, 535.1
Grandad, it ain‘t what you eat, it depends when you eat it-that‘s how muscles grow! Michael J. Rennie J. of Physiology 2001, 535.1

100
Energiebereitstellung
Abbau von Fettsäuren: -Oxidation Fette (Triacylglycerole) bilden die wichtigste Energiereserve des Organismus und sind vorwiegend im Fettgewebe gespeichert (v.a. im subkutanen Fettgewebe). Dort unterliegen sie einem ständigen Auf- und Abbau. Werden Fette gespalten (LIPOLYSE), entsteht Glycerol und freie Fettsäuren (FFS) [Die Lipolyse wird durch eine hormon-sensitive Lipase katalysiert, die einer komplexen hormonellen Kontrolle unterliegt]
 bilden die wichtigste Energiereserve des Organismus und sind vorwiegend im Fettgewebe gespeichert (v.a. im subkutanen Fettgewebe). Dort unterliegen sie einem ständigen Auf- und Abbau. Werden Fette gespalten (LIPOLYSE), entsteht Glycerol und freie Fettsäuren (FFS) [Die Lipolyse wird durch eine hormon-sensitive Lipase katalysiert, die einer komplexen hormonellen Kontrolle unterliegt]")
102
Energiebereitstellung
Abbau von Fettsäuren: -Oxidation Die Menge der vom Fettgewebe freigesetzten FFS ist dabei von der Aktivität der Lipase abhängig; das Enzym steuert den Plasmaspiegel der FFS Die vom Fettgewebe abgegebenen Fettsre. werden im Plasma unverestert transportiert: „free fatty acids“ (FFA) Nur die kurzkettigen FFS sind gelöst, die längeren, weniger wasserlöslichen FFS werden an Albumin gebunden.
 Nur die kurzkettigen FFS sind gelöst, die längeren, weniger wasserlöslichen FFS werden an Albumin gebunden.")
103
Energiebereitstellung
Abbau von Fettsäuren: -Oxidation Zum Verbrauch werden die FFS aus dem Plasma in die Zellen aufgenommen, wo sie zunächst proteingebunden vorliegen. Mit Ausnahme des Gehirns und der Erythrocyten sind alle Gewebe in der Lage, FFS durch -Oxidation abzubauen. Besonders intensiv ist der Stoffwechsel der FFS in der Leber – dort werden bei hohem FFS-Plasmaspiegel (zB. beim Hungern, Typ-1 Diabetes) v.a. Ketonkörper erzeugt.
 v.a. Ketonkörper erzeugt.")
104
Energiebereitstellung
Abbau von Fettsäuren: -Oxidation Nach der Aufnahme in die Zellen werden die FFS durch Überführung in ihr Co-Derivat aktiviert, es entsteht Acyl-CoA. Dafür werden 2 energiereiche Phosphatbindungen von ATP benötigt. Die Einschleusung in die Mitochondrien ermöglicht Carnitin, das als membrangängiger Carrier für die FFS fungiert.

105
Carnitin-Carrier

106
Energiebereitstellung
Abbau von Fettsäuren: -Oxidation Für FFS gibt es ein gruppenspezifisches Transport System, die aktivierten FFS (Acyl-CoA) müssen im Zytoplasma erst auf Carnitin übertragen werden, bevor sie von einem Carnitin-Carrier als Acyl-Carnitin im Tausch gegen freies Carnitin in den Matrix-Raum eingeschleust werden. Dort findet dann die Rückübertragung der FFS auf CoA statt
 müssen im Zytoplasma erst auf Carnitin übertragen werden, bevor sie von einem Carnitin-Carrier als Acyl-Carnitin im Tausch gegen freies Carnitin in den Matrix-Raum eingeschleust werden. Dort findet dann die Rückübertragung der FFS auf CoA statt.")
107
Energiebereitstellung
Abbau von Fettsäuren: -Oxidation Abbau der FFS findet in der Mitochondrien-Matrix statt und zwar durch einen oxidativen Reaktionszyklus, bei dem C2-Einheiten als Acetyl-CoA (‚aktivierte Essigsäure‘) abgespalten werden. Die schrittweise Abspaltung der Acetyl-Gruppen beginnt am Carboxy-Ende der aktivierten Fettsäuren jeweils zwischen C-2 (-Atom) und C-3 (-Atom) -Oxidation (räumlich und funktional ist diese mit dem Citrazyklus verbunden)
 abgespalten werden. Die schrittweise Abspaltung der Acetyl-Gruppen beginnt am Carboxy-Ende der aktivierten Fettsäuren jeweils zwischen C-2 (-Atom) und C-3 (-Atom) -Oxidation (räumlich und funktional ist diese mit dem Citrazyklus verbunden)")
108
Energiebereitstellung
Abbau von Fettsäuren: -Oxidation Exkurs: Fettsäuren Fettsäuren sind Carbonsäuren mit langer Kohlenwasserstoff-Kette Als Bausteine von Lipiden kommen sie in allen Organismen vor. FS sind vorwiegend mit Alkoholen verestert (z.B. Glycerol, Cholesterol, etc.) aber können auch unverestert auftreten (FFA) Die Zahl der C-Atome der längeren, natürlichen FS ist stets gerade (wegen der Biosynthese aus C2-Bausteinen)
 aber können auch unverestert auftreten (FFA) Die Zahl der C-Atome der längeren, natürlichen FS ist stets gerade (wegen der Biosynthese aus C2-Bausteinen)")
109
Energiebereitstellung
Abbau von Fettsäuren: -Oxidation Manche FS tragen Doppelbindungen (ungesättigte FS), diese tragen die erste DB meist zwischen C-9 und C-10. In mehrfach ungesättigten FS treten die anderen DB dann im Abstand von drei C-Atomen auf („isolierte DB„). FS mit konjugierten DB (Abstand von zwei C-Atomen) sind selten FS-Moleküle sind beweglich (aufgrund der freien Drehbarkeit der C-C Einfachbindung)
, diese tragen die erste DB meist zwischen C-9 und C-10. In mehrfach ungesättigten FS treten die anderen DB dann im Abstand von drei C-Atomen auf („isolierte DB„). FS mit konjugierten DB (Abstand von zwei C-Atomen) sind selten. FS-Moleküle sind beweglich (aufgrund der freien Drehbarkeit der C-C Einfachbindung)")
110
Aufbau aliphatischer Carbonsäuren

111
Struktur der Ölsäure

112
Energiebereitstellung
Abbau von Fettsäuren: -Oxidation Die Schmelzpunkte der FS steigen mit der Kettenlänge an, und fallen mit der Anzahl der DB ab Essentielle FS (mehrfach ungesättigte C-20 FS) werden vom Organismus für die Biosynthese von Eicosanoiden benötigt (die wichtigste essentielle FS Arachidonsäure, diese kann durch die beiden C-18 FS Linolsäure u. Linolensäure ersetzt werden)
 werden vom Organismus für die Biosynthese von Eicosanoiden benötigt (die wichtigste essentielle FS Arachidonsäure, diese kann durch die beiden C-18 FS Linolsäure u. Linolensäure ersetzt werden)")
113
Energiebereitstellung
Abbau von Fettsäuren: -Oxidation Eicosanoide sie dienen als second messenger einiger Hormone kontrollieren die Kontraktion der glatten Muskulatur (zB. Blutdruck, Bronchien…) greifen in die Ausschüttung von Zellprodukten ein beeinflussen den Knochenstoffwechsel, das autonome NS, das Immunsystem, die Zellwanderung und Zellaggregation sie sind wirkungsvolle Schmerzsignale für Nocizeptoren Eicosanoide wirken über Membranrezeptoren als ‚lokale Hormone‘ und weisen eine vielfältige biolog. Wirkung auf:
 greifen in die Ausschüttung von Zellprodukten ein. beeinflussen den Knochenstoffwechsel, das autonome NS, das Immunsystem, die Zellwanderung und Zellaggregation. sie sind wirkungsvolle Schmerzsignale für Nocizeptoren. Eicosanoide wirken über Membranrezeptoren als ‚lokale Hormone‘ und weisen eine vielfältige biolog. Wirkung auf:")
114
Energiebereitstellung
Abbau von Fettsäuren: -Oxidation Erster Schritt der -Oxidation ist die Dehydrierung von Acetyl-CoA wobei der Wasserstoff auf ein FAD-haltiges Protein übertragen wird das den Kontakt zur Atmungskette herstellt. Zweiter Schritt ist die Anlagerung von Wasser (Hydratisierung) an die DB der ungesättigten FS Dritter Schritt oxidiert die Hydroxy-Gruppe an C-3 zur Carbonyl-Gruppe (Dehydrierung) - Akzeptor für die Reduktionsäquivalente ist NAD+, das sie an die Atmungskette weiterreicht
 an die DB der ungesättigten FS. Dritter Schritt oxidiert die Hydroxy-Gruppe an C-3 zur Carbonyl-Gruppe (Dehydrierung) - Akzeptor für die Reduktionsäquivalente ist NAD+, das sie an die Atmungskette weiterreicht.")
115
Energiebereitstellung
Abbau von Fettsäuren: -Oxidation Vierter Schritt ist die thioklastische Spaltung – die aktivierte -Ketosäure wird durch eine Acyl-Transferase (-Ketothiolase) gespalten; dadurch entsteht Acetyl-CoA und eine aktivierte FS die gegenüber der ursprünglichen FS um eine C2-Einheit verkürzt ist Für den vollständigen Abbau langkettiger FS muß der Zyklus mehrfach durchlaufen werden (z.B. für Stearyl-CoA achtmal)
 gespalten; dadurch entsteht Acetyl-CoA und eine aktivierte FS die gegenüber der ursprünglichen FS um eine C2-Einheit verkürzt ist. Für den vollständigen Abbau langkettiger FS muß der Zyklus mehrfach durchlaufen werden (z.B. für Stearyl-CoA achtmal)")
116
Energiebereitstellung
Abbau von Fettsäuren: -Oxidation Das gebildete Acetyl-CoA wird auf Oxalacetat übertragen, dadurch entsteht Citrat, das den Citrat-Zyklus speist (oder in der Leber bei einem Überangebot von Acetyl-CoA der Synthese von Ketonkörpern dient )
")
117
Fettsäuren Abbau, -Oxidation

118
Energiebereitstellung
Abbau von Fettsäuren: -Oxidation Energiebilanz (am Bsp. der Palmitinsäure) Wird ein Molekül PS (16:0) vollständig zu 16 Molekülen CO2 oxidiert (wobei die -Oxidation in 7 Zyklen erfolgt) entstehen insgesamt 129 energiereiche DB je Molekül: Dies entspricht einer Energie von insgesamt 3935 kJ x mol-1
 Wird ein Molekül PS (16:0) vollständig zu 16 Molekülen CO2 oxidiert (wobei die -Oxidation in 7 Zyklen erfolgt) entstehen insgesamt 129 energiereiche DB je Molekül: Dies entspricht einer Energie von insgesamt 3935 kJ x mol-1.")
119
Energiebereitstellung
ATMUNGSKETTE Die Atmungskette katalysiert den Transport von Elektronen auf NADH oder reduziertem Ubichinon auf molekularen Sauerstoff Ein großer Teil der bei diesen Reaktionen gewonnen Energie wird zum Aufbau eines elektrochemischen Gradienten verwendet und dieser schließlich durch ATP-Synthase zur ATP-Bildung genutzt

120
Energiebereitstellung
ATMUNGSKETTE Die Elektronen werden auf verschiedenen Wegen in die Atmungskette eingeschleusst und im Rahmen der Oxidation an die verschiedenen Komplexe ‚weitergegeben‘, wobei durch den Elektronenfluss ein Protonengradient aufgebaut wird

121
Energiebereitstellung
ATMUNGSKETTE Anordnung Der Protonentransport durch einzelne Komplexe verläuft vektoriell von der Matrix in den Intramembranraum und erhöht dort die [H+] (der pH-Wert sinkt) - in intakten Mitochondrien erlaubt nur die ATP-Synthase den Rückfluss von Protonen in die Matrix (darauf beruht die regulatorisch wichtige energetische Kopplung von Elektronentransport und ATP-Bildung)
 - in intakten Mitochondrien erlaubt nur die ATP-Synthase den Rückfluss von Protonen in die Matrix (darauf beruht die regulatorisch wichtige energetische Kopplung von Elektronentransport und ATP-Bildung)")
122
Energiebereitstellung
ATMUNGSKETTE Die Oxidation von NADH findet an der Innenseite der Membran statt (im Matrixraum) wo auch Citrat-Zyklus und -Oxidation (die wichtigsten NADH-Lieferanten) lokalisiert sind. Auch die O2-Reduktion und die ATP-Bildung laufen an der Innenseite der Membran ab – das gebildete ATP wird schließlich im Antiport (mit Hilfe eines ADP/ATP Translokators) gegen ADP nach außen (Zytoplasma) transportiert
 wo auch Citrat-Zyklus und -Oxidation (die wichtigsten NADH-Lieferanten) lokalisiert sind. Auch die O2-Reduktion und die ATP-Bildung laufen an der Innenseite der Membran ab – das gebildete ATP wird schließlich im Antiport (mit Hilfe eines ADP/ATP Translokators) gegen ADP nach außen (Zytoplasma) transportiert.")
123
Aufbau der Atmungskette

124
Schematische Darstellung des FS-Abbaus
Karl-Franzens: Schematische Darstellung des FS-Abbaus

125
Energiebereitstellung
Steuerung des FS-Abbaus Der Spiegel der FFS steht unter der Kontrolle jener Hormone, die die Lipolyse im Fettgewebe steuern. Aus dem Plasmapool der FFS versorgen sich die Gewebe mit (1) FS (soweit sie die FS nicht selbst durch eigene Synthese – Lipogenese) bilden (2). Nach der Aktivierung zu Acyl-CoA (3) werden die intrazellulären FFS entweder gespeichert oder abgebaut
 FS (soweit sie die FS nicht selbst durch eigene Synthese – Lipogenese) bilden (2). Nach der Aktivierung zu Acyl-CoA (3) werden die intrazellulären FFS entweder gespeichert oder abgebaut.")
126
Energiebereitstellung
Steuerung des FS-Abbaus Speicherung Die Veresterung der aktivierten FS (Acyl-CoA) mit Glycerol (4) führt zu Phospholipiden, die als Membranbausteine benötigt werden, und zu Triacylglycerolen (Fett) die als Energiereserve gespeichert werden (5). Die Leber kann diese Produkte auch als Lipoproteine an das Plasma abgeben (6) [wenn reichlich FFS vorhanden sind]
 mit Glycerol (4) führt zu Phospholipiden, die als Membranbausteine benötigt werden, und zu Triacylglycerolen (Fett) die als Energiereserve gespeichert werden (5). Die Leber kann diese Produkte auch als Lipoproteine an das Plasma abgeben (6) [wenn reichlich FFS vorhanden sind]")
127
Energiebereitstellung
Steuerung des FS-Abbaus Abbau Der Abbau der FS erfolgt in den Mitochondrien. Die FS werden mittels des Carnitin-Shuttles aus dem Zytoplasma in die Mitochondrien transportiert (7). Sie werden dort oxidativ durch Zusammenwirken von -Oxidation, Citrat-Zyklus und Atmungskette zu CO2 abgebaut (8+9), dabei wird in erheblichem Umfang ATP gewonnen. In den Mitochondrien der Leber können Ketonkörper gebildet werden (10) (Ketogenese)
. Sie werden dort oxidativ durch Zusammenwirken von -Oxidation, Citrat-Zyklus und Atmungskette zu CO2 abgebaut (8+9), dabei wird in erheblichem Umfang ATP gewonnen. In den Mitochondrien der Leber können Ketonkörper gebildet werden (10) (Ketogenese)")
128
Energiebereitstellung
Steuerung des FS-Abbaus Wer bestimmt die Geschwindigkeit des FS-Abbaus? Der Abbau ist abhängig von Transport der FS in die Mitochondrien - dessen erster Schritt ist die Übertragung der FS von CoA auf Carnitin durch eine Transferase (Carnitin-O-Palmitoyl-Transferase) Dieses in der äußeren Mitochondrienmembran lokalisierte Enzym ist im Hungerzustand besonders aktiv, wenn der Plasmaspiegel von FFS und Glukagon hoch ist
 Dieses in der äußeren Mitochondrienmembran lokalisierte Enzym ist im Hungerzustand besonders aktiv, wenn der Plasmaspiegel von FFS und Glukagon hoch ist.")
129
Energiebereitstellung
Steuerung des FS-Abbaus Im Sättigungszustand wird die Transferase durch Malonyl-CoA gehemmt (wird im Sättigungszustand angehäuft). Durch den verringerten Import von FS in die Mitochondrien ist dann die -Oxidation gedrosselt. Gleichzeitig ist die Lipogenese gesteigert, da ihr Schlüsselenzym, die Acetyl-CoA-Carboxylase stimuliert ist.
. Durch den verringerten Import von FS in die Mitochondrien ist dann die -Oxidation gedrosselt. Gleichzeitig ist die Lipogenese gesteigert, da ihr Schlüsselenzym, die Acetyl-CoA-Carboxylase stimuliert ist.")
130
Energiebereitstellung
Steuerung des FS-Abbaus Wie kann ein vermehrter Abbau von FS noch reguliert werden? 1. Training im Hungerzustand (postresorbtiven Phase od auf nüchternen Magen (‚Morgensport‘) 2. Ernährungsmodifikationen (Reduktion des CHO-Anteils, Erhöhung des Anteils an Fetten (v.a. mehrfach ungesättigte) und Proteinen bzw generell eine Verringerung der Gesamtkalorien- zufuhr
 2. Ernährungsmodifikationen (Reduktion des. CHO-Anteils, Erhöhung des Anteils an Fetten. (v.a. mehrfach ungesättigte) und Proteinen bzw. generell eine Verringerung der Gesamtkalorien- zufuhr.")
131
Energiebereitstellung
Steuerung des FS-Abbaus 3. Ernährung nach dem Glykämischen Index (niedrige Insulinspiegel infolge längerdauernder Spaltprozesse) 4.Trainingsreihenfolge beachten (zuerst die kraftorientierten Anteile, dann die ausdauerorientierten Anteile) 5.Beachtung der Tageszeiten für das Training (endokrinen Rhythmen) und der Schlafphase (verschiedene AS fördern die Lipolyse über hormonelle Regulationsmechanismen)
 4.Trainingsreihenfolge beachten (zuerst die kraftorientierten Anteile, dann die ausdauerorientierten Anteile) 5.Beachtung der Tageszeiten für das Training (endokrinen Rhythmen) und der Schlafphase (verschiedene AS fördern die Lipolyse über hormonelle Regulationsmechanismen)")
132
Energiebereitstellung
Zur Erinnerung Die ATP-Produktion muß ständig dem wechselnden Bedarf angepasst werden Der Wiederaufbau des ATP aus ADP erfolgt auf 3 Wegen Aus KP = anaerob alaktazide Resynthese Über die anaerobe Oxidation = anaerob laktazide Resynthese Über die aerobe Oxidation = aerobe Resynthese

133
Energiebereitstellung
Der kleinere Energiespeicher (ATP) beliefert DIREKT die energiebenötigenden Reaktionen Der größere Energiespeicher (KP) füllt den ATP-Speicher wieder auf Bei einem ausreichenden Sauerstoffangebot (aerobe Oxidation) erfolgt der Abbau von KH und Fetten (bzw. Proteinen) SCHRITTWEISE in enzymgesteuerten Stufen zu CO2 und H2O (energie- freie Endprodukte)
 beliefert DIREKT die energiebenötigenden Reaktionen. Der größere Energiespeicher (KP) füllt den ATP-Speicher wieder auf. Bei einem ausreichenden Sauerstoffangebot. (aerobe Oxidation) erfolgt der Abbau von. KH und Fetten (bzw. Proteinen) SCHRITTWEISE in. enzymgesteuerten Stufen zu CO2 und H2O (energie- freie Endprodukte)")
134
Energiebereitstellung
Aerobe Oxidation: Glykogenolyse (Glykogenabbau) Glykolyse (Glukoseabbau) Bildung der aktivierten Essigsäure (Azetyl-CoA) damit das gebildete Pyruvat für den weiteren Abbau verwendet werden kann Trikarbonsäure- oder Citrazyklus (auch Krebszyklus genannt) Atmungskette (oder Elektronentransportkette)
 Glykolyse (Glukoseabbau) Bildung der aktivierten Essigsäure (Azetyl-CoA) damit das gebildete Pyruvat für den weiteren Abbau verwendet werden kann. Trikarbonsäure- oder Citrazyklus (auch Krebszyklus genannt) Atmungskette (oder Elektronentransportkette)")
135
Energiebereitstellung
Wenn das Sauerstoffangebot nicht ausreicht, dann wird der Weg der anaeroben Glykolyse beschritten [dann, wenn der momentane Energiebedarf nicht durch aerobe Oxidation gedeckt werden kann] Die maximale aerobe Oxidation wird begrenzt: O2-Menge, die der Muskelzelle/Zeiteinheit zugeführt werden kann Menge der Enzyme, die die einzelnen Reaktionen bei der Oxidation katalysieren Größe der Nährstoffdepots in der Muskelzelle sowie deren Wiederauffüllung

136
Energiebereitstellung
Anaerobe Energiebereitstellung In Abwesenheit von Sauerstoff (zeitabhängige Transportprozeße an der inneren Mitochondrien-membran – z.T. TRAINIERBAR ! – durch Zunahme der Oberfläche und tlw. genetisch bestimmt aufgrund der Muskelfaserverteilung) kann in den Mitochondrien NADH nicht mehr reoxidiert werden; damit kommt außer der ATP-Bildung auch der Citrat-Zyklus (und damit der FS-und AS-Abbau) zum Erliegen Ort des Geschehens: die Enzyme befinden sich im Sarkoplasma (in unmittelbarer Nähe der Myofibrillen)
 kann in den Mitochondrien NADH nicht mehr reoxidiert werden; damit kommt außer der ATP-Bildung auch der Citrat-Zyklus (und damit der FS-und AS-Abbau) zum Erliegen. Ort des Geschehens: die Enzyme befinden sich im Sarkoplasma (in unmittelbarer Nähe der Myofibrillen)")
137
Energiebereitstellung
Anaerobe Energiebereitstellung Als Energiesubstrat bleibt dann nur mehr Glukose übrig [der Abbau im Rahmen der Glykolyse zu Pyruvat (ionisierte Form der Brenztraubensäure) liefert 2 mol ATP] Wenn dieser Prozess kontinuierlich weiter laufen soll (wenn also bei hoher Intensität die Leistung aufrecht erhalten werden muß), dann muss das in der Glykolyse gebildete NADH außerhalb der Mitochondrien regeneriert werden – dazu wird Pyruvat zu Laktat reduziert
 liefert 2 mol ATP] Wenn dieser Prozess kontinuierlich weiter laufen soll (wenn also bei hoher Intensität die Leistung aufrecht erhalten werden muß), dann muss das in der Glykolyse gebildete NADH außerhalb der Mitochondrien regeneriert werden – dazu wird Pyruvat zu Laktat reduziert.")
138
Energiebereitstellung
Anaerobe Energiebereitstellung Warum ist das so? Wird die Glykolyse durch eine starke Steigerung des Energiebedarfs gefordert, so fällt mehr Brenztraubensäure an, als oxidativ verarbeitet werden kann; dadurch wird die Glykolyse verlangsamt und damit die anaerobe Energiebereitstellung gefährdet [Ursache: neben Brenztraubensäure häuft sich NADH+H+ (reduzierte Form) an – dieses muss aber als NAD+ vorliegen (in oxidierter Form) um als H+-Akzeptor fungieren zu können]
 an – dieses muss aber als NAD+ vorliegen (in oxidierter Form) um als H+-Akzeptor fungieren zu können]")
139
Energiebereitstellung
Anaerobe Energiebereitstellung Ausweg: Da die Atmungskette nicht so rasch das anfallende NADH+H+ oxidieren kann wird Wasserstoff des NADH+H+ auf Pyruvat übertragen, wobei Laktat und das benötigte NAD+ entsteht De facto ist die AN-Glykolyse ein Stoffwechselweg, der unabhängig vom aktuellen Zustand der aeroben Oxidation ablaufen kann; Probleme macht die Tatsache, dass die [H+] proportional zur Laktatkonzentration ansteigt (unter physiolog. Bedingungen ist Laktat zu über 95% dissoziiert)
")
141
Energiebereitstellung
Anaerobe Energiebereitstellung Das Enzym das die Reduktion zu Laktat katalysiert ist die Laktatdehydrogenase (2 verschiedene LDH Untereinheiten: H- bzw. M-Form, und 5 Isoenzyme der LDH) In der Herz-Muskulatur findet sich vorwiegend Tetramere der H-Form, in der LDH der Leber und der Skelettmuskulatur vorwiegend M-Monomere
 In der Herz-Muskulatur findet sich vorwiegend Tetramere der H-Form, in der LDH der Leber und der Skelettmuskulatur vorwiegend M-Monomere.")
142
Energiebereitstellung
Anaerobe Energiebereitstellung Die LDH katalysiert die Übertragung von Redox-Äquivalenten von Laktat auf NAD+ oder von NADH auf Pyruvat Laktat + NAD+ Pyruvat + NADH + H+ Die LDH katalysiert die Reaktion in beide Richtungen, hat aber keinen Einfluss auf die Lage des chem. Gleichgewichts; das Gleichgewicht der Reaktion liegt aber stark auf der Seite der Laktat-Bildung; bei hohen Konzentrationen von Laktat und NAD+ ist aber auch die Oxidation von Laktat zu Pyruvat möglich

143
Energiebereitstellung
Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Das wichtigste regulierende Enzym der Glykolyse ist die PFK; sie wird durch ADP, AMP, und F-6-P als Substrat aktiviert und durch ATP und H+-Ionen gehemmt. Sinkt der pH-Wert auf z.B. 6.7 ab, so beträgt die Aktivität der PFK (und damit die Flußrate) nur mehr ~ 50% des Maximums; bei einem pH-Wert von ~ 6.3 ist sie komplett gehemmt (die ATP-Resyntheserate aus der Glykolyse geht gegen Null)
 nur mehr ~ 50% des Maximums; bei einem pH-Wert von ~ 6.3 ist sie komplett gehemmt (die ATP-Resyntheserate aus der Glykolyse geht gegen Null)")
144
Energiebereitstellung
Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Die pH-bedingte Hemmung der Glykolyse schützt vor der zunehmenden Aktivierung von Lysosomen [ µm große Organellen, die dem Abbau von Zellbestandteilen dienen] – damit wird eine „Selbstverdauung“ der Zelle verhindert Die Übersäuerung der Zelle kann verzögert werden durch: 1. die in der Zelle vorhandenen Puffersysteme 2. Die Entfernung des Laktats

145
Energiebereitstellung
Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Das anfallende Laktat wird bereits während der Arbeit weiterverwertet (v.a. bei ausreichend durchbluteter Muskulatur); in der Arbeitsmuskulatur (hptsl. in den roten MF und in den weniger stark belasteten Faseranteilen, die sich außerhalb der Belastungsachsen befinden), in der nicht-arbeitenden Muskulatur, in der Herzmuskulatur (der Herzmuskel ist aufgrund seiner aeroben Leistungsfähigkeit in der Lage, einen großen Prozentsatz seines Energiebedarfs über Laktat abzudecken)
; in der Arbeitsmuskulatur (hptsl. in den roten MF und in den weniger stark belasteten Faseranteilen, die sich außerhalb der Belastungsachsen befinden), in der nicht-arbeitenden Muskulatur, in der Herzmuskulatur (der Herzmuskel ist aufgrund seiner aeroben Leistungsfähigkeit in der Lage, einen großen Prozentsatz seines Energiebedarfs über Laktat abzudecken)")
146
Energiebereitstellung
Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Ein Teil des Laktats wird in der Leber wieder zu Glukose aufgebaut (Glukoneogenese) [Dabei wird das im Blut zirkulierende Laktat zur Leber transportiert und im Rahmen des CORI-Zyklus zu Glukose umgewandelt und wieder zum Verbraucher zurücktransportiert]
 [Dabei wird das im Blut zirkulierende Laktat zur Leber transportiert und im Rahmen des CORI-Zyklus zu Glukose umgewandelt und wieder zum Verbraucher zurücktransportiert]")
147
Energiebereitstellung
Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Der messbare Laktatanstieg im Blut wäre noch größer, wenn nicht ein Teil des anfallenden Pyruvats durch Aminierung (Substitutionsreaktion) in die AS Alanin übergeführt würde (Alanin wird in der Leber - aber nicht im Muskel - zu Glukose und Glykogen aufgebaut) – Alanin-Zyklus (In der Erholungsphase kann das durch anerobe Belastung gebildete Laktat im Muskel zu ~ 50% über die Glukoneogenese wieder zu Glukose und Glykogen umgewandelt werden)
 in die AS Alanin übergeführt würde (Alanin wird in der Leber - aber nicht im Muskel - zu Glukose und Glykogen aufgebaut) – Alanin-Zyklus. (In der Erholungsphase kann das durch anerobe Belastung gebildete Laktat im Muskel zu ~ 50% über die Glukoneogenese wieder zu Glukose und Glykogen umgewandelt werden)")
148
Energiebereitstellung
Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Der Alanin-Zyklus wird durch den Abbau von Proteinen gespeist - die dabei anfallenden AS werden durch Transaminierung in 2-Oxosäuren umgewandelt, die großteils in den Citratzyklus wandern; dabei werden ihre Amino-Gruppen auf Pyruvat übertragen, das in Alanin übergeht (der Alanin-Zyklus dient glz. dem Transport von Glukose-Vorstufen und Stickstoff zur Leber – in der Leber wird N zu Harnstoff umgesetzt)
")
150
Energiebereitstellung
Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Nach intensiven Belastungen (z.B. 400m-Lauf) steigt das Blutlaktat noch weiter an, und erreicht in Abhängigkeit von Belastungsdauer und [LA] ein Maximum nach 1-10min. Erholungszeit – die Ursache dafür ist der Konzentrationsgradient zwischen Muskel- und Blutlaktatkonzentration Für die Erfassung der LA-Elimination wird die Halbwertszeit (Zeit, nach der das LA auf die Hälfte des max. Wertes abgebaut ist) angegeben (diese hängt von der Höhe des maximal erreichten LA-Wertes ab!)
 steigt das Blutlaktat noch weiter an, und erreicht in Abhängigkeit von Belastungsdauer und [LA] ein Maximum nach 1-10min. Erholungszeit – die Ursache dafür ist der Konzentrationsgradient zwischen Muskel- und Blutlaktatkonzentration. Für die Erfassung der LA-Elimination wird die Halbwertszeit (Zeit, nach der das LA auf die Hälfte des max. Wertes abgebaut ist) angegeben (diese hängt von der Höhe des maximal erreichten LA-Wertes ab!)")
151
Laktatbildung und LA Verhalten im Muskel und im Blut
(de Marées, H.: Sportphysiologie)
")
152
Energiebereitstellung
Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Im Mittel geht man von ½-T von 15min. bei einer [LA] von 10mmol aus; bei sehr hohen [LA] kann ½-T 25min. überschreiten Die Ursache für diese konzentrationsabhängige ½-T wird in der Existenz eines ‚Laktat-Shuttle‘ Systems gesehen; dabei dürfte das LA mittels spezifischer Proteine durch Zellmembranen transportiert werden; die Transportrate ist dabei eine nichtlineare Funktion der [LA]

153
Laktatverhalten im Blut (1/2-T) bei unterschiedlichen
Karl-Franzens: Laktatverhalten im Blut (1/2-T) bei unterschiedlichen LA-Ausgangswerten (de Marées, H.: Sportphysiologie)
 bei unterschiedlichen. LA-Ausgangswerten (de Marées, H.: Sportphysiologie)")
154
Energiebereitstellung
Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Aktive Erholung (leichte Belastung) verkürzt die ½-T der LA-Elimination! Maximale Eliminationsraten erreicht man bei Belastungen von 40-50% der maximalen VO2 [durch den erhöhten Muskelstoffwechsel wird Laktat vermehrt in Brenztraubensäure übergeführt und in weiterer Folge als direkt energielieferndes Substrat mittels aerober Oxidation verstoffwechselt; d.h. Laktat ist energetisch ‚wertvoll‘
 verkürzt die ½-T der LA-Elimination! Maximale Eliminationsraten erreicht man bei Belastungen von 40-50% der maximalen VO2 [durch den erhöhten Muskelstoffwechsel wird Laktat vermehrt in Brenztraubensäure übergeführt und in weiterer Folge als direkt energielieferndes Substrat mittels aerober Oxidation verstoffwechselt; d.h. Laktat ist energetisch ‚wertvoll‘")
155
Einfluß von passiver Erholung und aktiver Erholung auf das Blutlaktat nach einer Belastung (de Marées, H.: Sportphysiologie)
")
156
Energiebereitstellung
Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Laktat wird im Stoffwechsel immer gebildet (Ruhelaktatwerte von ~ mmol/l), und diese Bildung hängt u.a. auch von der Ernährung ab Die LA-Elimination hängt von der Höhe des aeroben Umsatzes und von der [LA] ab. Je höher der aerobe Stoffwechsel und je höher die [LA], desto größer die Elimination Die jeweils aktuelle [LA] im Muskel, im Blut und in anderen Körperflüssigkeiten ist das Resultat von LA-bildung,-diffusion,-transport, und -elimination
, und diese Bildung hängt u.a. auch von der Ernährung ab. Die LA-Elimination hängt von der Höhe des aeroben Umsatzes und von der [LA] ab. Je höher der aerobe Stoffwechsel und je höher die [LA], desto größer die Elimination. Die jeweils aktuelle [LA] im Muskel, im Blut und in. anderen Körperflüssigkeiten ist das Resultat von. LA-bildung,-diffusion,-transport, und -elimination.")
157
Energiebereitstellung
Anaerobe Energiebereitstellung Im Vergleich zum Gesamtgewinn von 31 mol ATP (wenn 1 mol Glukose abgebaut wird) bei der aeroben Oxidation, ergeben sich für die anaerobe Glykolyse 3 mol ATP/mol Glukose (~ 10%) Da die maximale Umsatzrate der Glukose jedoch 25fach höher ist als die von Citratzyklus und Atmungskette (hier geht man von den Werten für Untrainierte aus), resultiert eine etwa doppelt so große ATP-Resynthese/Zeiteinheit aus der anaeroben Glykolyse
 bei der aeroben Oxidation, ergeben sich für die anaerobe Glykolyse 3 mol ATP/mol Glukose (~ 10%) Da die maximale Umsatzrate der Glukose jedoch 25fach höher ist als die von Citratzyklus und Atmungskette (hier geht man von den Werten für Untrainierte aus), resultiert eine etwa doppelt so große ATP-Resynthese/Zeiteinheit aus der anaeroben Glykolyse.")
158
Vorteile/Nachteile der aeroben und anaeroben Energiebereitstellung
Aerobe EB Anaerobe EB 31 mol ATP/mol Glukose 3 mol ATP/mol Glukose Keine LA-Bildung LA-Bildung (-) langsame Energie-bereitstellung (+) schnelle Energie-bereitstellung (-) freigesetzte Energ.-menge/Zeiteinheit ist relativ klein (+) freigesetzte Energ.menge/Zeiteinheit ist relativ groß
 langsame Energie-bereitstellung. (+) schnelle Energie-bereitstellung. (-) freigesetzte Energ.-menge/Zeiteinheit ist relativ klein. (+) freigesetzte Energ.menge/Zeiteinheit ist relativ groß.")
159
Vorteile/Nachteile der aeroben und anaeroben Energiebereitstellung
Aerobe EB Anaerobe EB (+) bereitgestellte Energiemenge ist relativ groß (-) die Gesamtenergiemenge ist relativ klein Sportliche Tätigkeiten können relativ lange durchgehalten werden, wenn die Intensität nicht zu hoch ist (1000m Lauf) Sportliche Tätigkeiten können mit hoher Intensität geleistet werden, aber nur kurzzeitig (400m Lauf)
 bereitgestellte Energiemenge ist relativ groß. (-) die Gesamtenergiemenge ist relativ klein. Sportliche Tätigkeiten können relativ lange durchgehalten werden, wenn die Intensität nicht zu hoch ist (1000m Lauf) Sportliche Tätigkeiten können mit hoher Intensität geleistet werden, aber nur kurzzeitig (400m Lauf)")
160
Laktatverhalten und Anteile der Energie-gewinnung bei unterschiedlichen
(Markworth, P. SPORTMEDIZIN) Belastungsintensitäten
 Belastungsintensitäten.")
161
Anteil der energiebereitstellenden Systeme im zeitl
Anteil der energiebereitstellenden Systeme im zeitl.Verlauf einer körperlichen Belastung (Markworth, P. SPORTMEDIZIN)
")
162
Energiebereitstellung
1. HS der Thermodynamik: Die im Muskel umgesetzte (chemische) Energie muß gleich der Summe aus mechanischer Energie (Muskelarbeit) und Wärmeproduktion sein (Energieerhaltungssatz) Wenn, wie im Fall einer isometrischen Kontraktion keine physikalisch messbare Muskelarbeit erfolgt, wird im Muskel chemische Energie in Wärme transformiert; diese ist der Haltezeit und der aufrecht erhaltenden Muskelspannung proportional [bei der Isometrie sind die Myosinquerbrücken in ständiger zyklischer Ruder-tätigkeit wobei unter ATP-Spaltung und Wärmeproduktion eine ‚innere‘ Haltearbeit verrichtet wird]
 Energie muß gleich der Summe aus mechanischer Energie (Muskelarbeit) und Wärmeproduktion sein (Energieerhaltungssatz) Wenn, wie im Fall einer isometrischen Kontraktion keine physikalisch messbare Muskelarbeit erfolgt, wird im Muskel chemische Energie in Wärme transformiert; diese ist der Haltezeit und der aufrecht erhaltenden Muskelspannung proportional [bei der Isometrie sind die Myosinquerbrücken in ständiger zyklischer Ruder-tätigkeit wobei unter ATP-Spaltung und Wärmeproduktion eine ‚innere‘ Haltearbeit verrichtet wird]")
163
Energiebereitstellung
Wirkungsgrad: 1 Mol ATP ~ 60kJ Energie (Hydrolyse) Es werden allerdings nur etwa 40-50% dieser Energie in mechanische Energie bzw. Arbeit umgesetzt (der Rest geht als Wärme ‚verloren‘) Der mechanische Nutzeffekt (Wirkungsgrad) liegt aber de facto nur bei ~ 20-30% - während und nach der Kontraktion laufen Erholungsprozesse (Tätigkeit von Ionenpumpen, oxidative Regeneration von ATP) die für eine beträchtliche Wärmebildg. sorgen [je höher die Arbeitsleistung, desto höher die Wärmebildg.]
 Es werden allerdings nur etwa 40-50% dieser Energie in mechanische Energie bzw. Arbeit umgesetzt (der Rest geht als Wärme ‚verloren‘) Der mechanische Nutzeffekt (Wirkungsgrad) liegt aber de facto nur bei ~ 20-30% - während und nach der Kontraktion laufen Erholungsprozesse (Tätigkeit von Ionenpumpen, oxidative Regeneration von ATP) die für eine beträchtliche Wärmebildg. sorgen [je höher die Arbeitsleistung, desto höher die Wärmebildg.]")
164
Proteine (als Aminosäuren)
Energiebereitstellung Was steht als ‚Brennstoff‘ zur Verfügung? Kohlenhydrate (gespeichert in der Muskulatur und in der Leber bzw im Blut) Fette (als Depotfette-subkutan bzw viszeral und im Blut-aber auch intramuskulär) Proteine (als Aminosäuren) Der O2-Verbrauch beim Abbau der Brennstoffe hängt dabei von der Muskelleistung ab !
 Fette (als Depotfette-subkutan bzw viszeral und im Blut-aber auch intramuskulär) Proteine (als Aminosäuren) Der O2-Verbrauch beim Abbau der Brennstoffe. hängt dabei. von der Muskelleistung ab !")
165
Energiebereitstellung
ATP-Aufbau und Formen der ATP-Bildung Das Coenzym ATP stellt die wichtigste Speicherform chemischer Energie dar. Die bei der Spaltung von ATP gewonnene Energie dient dazu, durch energetische Kopplung Vorgänge wie Biosynthesen, Transportprozesse und Bewegungen zu ermöglichen Struktur: Im ATP ist eine Kette von 3 Phosphatresten mit der 5‘-OH-Gruppe von Adenosin verknüpft. Die Phosphatreste werden als , , und gekennzeichnet

166
Karl-Franzens:

167
Energiebereitstellung
ATP-Aufbau und ATP-Bildung Formen der ATP-Bildung Im Laufe der Evolution haben sich 2 wichtige Formen der ATP-Synthese herausgebildet, die in ähnlicher Weise in allen Zellen stattfinden (1) Die wichtigere Form nutzt die in elektrochemischen Gradienten gespeicherte Energie zur Bildung von ATP aus ADP und Pi Die Energie zum Aufbau solcher Gradienten wird meist durch Redoxprozesse aufgebracht (also die oxidative Phosphorylierung) [H+transportierende ATP-Synthasen nutzen dann die Energie des Gradienten zur ATP-Bildung]
 Die wichtigere Form nutzt die in elektrochemischen Gradienten gespeicherte Energie zur Bildung von ATP aus ADP und Pi. Die Energie zum Aufbau solcher Gradienten wird meist durch Redoxprozesse aufgebracht (also die oxidative Phosphorylierung) [H+transportierende ATP-Synthasen nutzen dann die Energie des Gradienten zur ATP-Bildung]")
168
Energiebereitstellung
ATP-Aufbau und ATP-Bildung (2) Die zweite, entwicklungsgeschichtlich ältere Form der ATP-Bildung findet auch unter anaeroben Bedingungen statt Diese beruht auf der Kopplung der ATP-Bildung an die Hydrolyse energiereicher Metabolite Als Beispiel ist hier die ATP-Bildung aus Kreatinphosphat zu nennen, einer Verbindung, die der Muskulatur als Energiereserve dient
 Die zweite, entwicklungsgeschichtlich ältere Form der ATP-Bildung findet auch unter anaeroben Bedingungen statt. Diese beruht auf der Kopplung der ATP-Bildung an die Hydrolyse energiereicher Metabolite. Als Beispiel ist hier die ATP-Bildung aus Kreatinphosphat zu nennen, einer Verbindung, die der Muskulatur als Energiereserve dient.")
169
Energiebereitstellung
ATP-Aufbau und ATP-Bildung ATP + H2O ADP + Pi (ATPase) Diese Rkt ist wichtig für die Energetik der Muskelkontraktion ATP + H2O AMP + PPi Diese Rkt ist v.a. für die Energie-lieferung bei Biosynthesereaktionen; dabei können auch beide Phosphatgruppen abgespalten werden und es entsteht AMP u. Pyrophosphorsäure bzw. Pyrophosphat (PPi)
 Diese Rkt ist wichtig für die Energetik der Muskelkontraktion. ATP + H2O AMP + PPi. Diese Rkt ist v.a. für die Energie-lieferung bei Biosynthesereaktionen; dabei können auch beide Phosphatgruppen abgespalten werden und es entsteht AMP u. Pyrophosphorsäure bzw. Pyrophosphat (PPi)")
170
Energiebereitstellung
ATP-Aufbau und ATP-Bildung Kreatinphosphat Durch die Muskelkontraktion wird die im Muskel vorhandene ATP-Menge in kürzester Zeit aufgebraucht (1-3 ? Sekunden bzw. max. 2-3 Kontraktionen); d.h. die Nachlieferung von ATP muß sehr rasch und effizient erfolgen KrP + ADP Kr + ATP (Lohmann Reaktion) Das Enzym, dass diese Reaktion katalysiert ist die Kreatinphosphokinase
; d.h. die Nachlieferung von ATP muß sehr rasch und effizient erfolgen. KrP + ADP Kr + ATP (Lohmann Reaktion) Das Enzym, dass diese Reaktion katalysiert ist die Kreatinphosphokinase.")
171
Energiebereitstellung
ATP-Aufbau und ATP-Bildung Das Enzym KrP-Kinase ist so aktiv, dass bereits während der Kontraktionsphase das angefallene ADP zu ATP aufgebaut werden kann. Im Normallfall bleibt daher auch die ATP-Konzentration bis zur völligen Ausschöpfung der KrP Speicher weitestgehend konstant Die Enzymaktivität ist weitgehend der Maximalgeschwindigkeit des ATP-Verbrauchs angepasst (~ 10 mal höher im Skelettmuskel als im Herzmuskel)
")
172
Energiebereitstellung
ATP-Aufbau und ATP-Bildung Ohne Nachlieferung aus dem aneroben oder aeroben Stoffwechsel kann der KrP-Speicher einen Arbeitsbetrag von ~ 900 J/kg Muskulatur liefern (kurzzeitige Höchstleistungen, z.B. 100m Lauf) Die „Flußrate“ (µmol x g-1 x s-1) [Geschwindigkeit des Abbaus und ~ ‚Einsatzzeitpunkt‘ der Verfügbarkeit]
 Die „Flußrate (µmol x g-1 x s-1) [Geschwindigkeit des Abbaus und ~ ‚Einsatzzeitpunkt‘ der Verfügbarkeit]")
173
Energiebereitstellung
ATP-Aufbau und ATP-Bildung Geschwindigkeit ATP, KrP Anaerobe Glykolyse Aerobe FS Oxidation

174
Energiebereitstellung
Brennwert und Energiespeicher ATP 1.5 kcal KrP 5 kcal Glucose (Blut) 20 kcal Leberglykogen 400 kcal Muskelglykogen 2000 kcal Freie Fettsre. u. Triglyceride 90 kcal Triglyceride (Blut) 2500 kcal Speicherfettdepots von kcal bis ?
 20 kcal. Leberglykogen 400 kcal. Muskelglykogen 2000 kcal. Freie Fettsre. u. Triglyceride 90 kcal. Triglyceride (Blut) 2500 kcal. Speicherfettdepots von kcal bis")
175
Energiebereitstellung
Energiequellen im Muskel Energiequelle Gehalt (µMol/g Muskel) Energ.-lief.Rkt ATP ATP -> ADP + P Kreatinphosphat KrP+ADP->ATP+Kr Gluk-Einheiten anaerob, über Pyr. im Glykogen zu Laktat(Glykolyse) aerob, über Pyr. zu CO2 u.H2O Triglyceride Oxidation zu CO u. H2O
 Energ.-lief.Rkt. ATP 5 ATP -> ADP + P. Kreatinphosphat 11 KrP+ADP->ATP+Kr. Gluk-Einheiten anaerob, über Pyr. im Glykogen 84 zu Laktat(Glykolyse) aerob, über Pyr. zu CO2 u.H2O. Triglyceride 10 Oxidation zu CO2 u. H2O.")
176
Energiebereitstellung
Aerober Energiestoffwechsel Ist die muskuläre Tätigkeit über einen längerdauernden Zeitraum aufrechtzuerhalten, so erfolgt der Wiederaufbau des ATP aerob (oxidative Phosphorylierung). Die zur Synthese notwendige Energie stammt dabei aus dem Abbau von Kohlehydraten und Fetten – dieses System befindet sich im Fließgleichgewicht, wenn die Geschwindigkeit der ATP-Neubildung gerade so groß ist wie die Geschwindigkeit des ATP-Verbrauchs [Spiegel des IZ ATP und des Kreatinphosphats bleiben dabei auf einem konstanten Niveau]
. Die zur Synthese notwendige Energie stammt dabei aus dem Abbau von Kohlehydraten und Fetten – dieses System befindet sich im Fließgleichgewicht, wenn die Geschwindigkeit der ATP-Neubildung gerade so groß ist wie die Geschwindigkeit des ATP-Verbrauchs. [Spiegel des IZ ATP und des Kreatinphosphats bleiben dabei auf einem konstanten Niveau]")
177
Energiebereitstellung
Aerober Energiestoffwechsel Die aerobe Glykolyse erfolgt in den Mitochondrien der Zellen (bakteriengroße Organellen, die ~ 25% des Zellvolumens einnehmen) – Mitochondrien sind die Hauptverbraucher von Sauerstoff; ein Mangel an O2 (z.B. Herzinfarkt, Arteriosklerose) wirkt sich besonders auf die Mitochondrien des betroffenen Gewebes aus. EXKURS: Aufbau der Mitochondrien Mitochondrien sind von 2 Membranen umschlossen-einer glatten äusseren und einer gefalteten inneren Membran (Cristae; große Oberfläche!) die den Matrix-Raum umschließt
 – Mitochondrien sind die Hauptverbraucher von Sauerstoff; ein Mangel an O2 (z.B. Herzinfarkt, Arteriosklerose) wirkt sich besonders auf die Mitochondrien des betroffenen Gewebes aus. EXKURS: Aufbau der Mitochondrien. Mitochondrien sind von 2 Membranen umschlossen-einer glatten äusseren und einer gefalteten inneren Membran (Cristae; große Oberfläche!) die den Matrix-Raum umschließt.")
178
Energiebereitstellung
EXKURS (Aufbau der Mitochondrien) Anzahl und Gestalt der Mitochondrien sowie die Zahl der Cristae kann von Zelltyp zu Zelltyp unterschiedlich sein. Gewebe mit intensivem oxidativen Stoffwechsel besitzen Mitochondrien mit vielen Cristae-innerhalb eines Gewebes kann die Gestalt der Mitochondrien mit ihrem Funktionszustand variieren Die Membranen der Mitochondrien enthalten integrale Proteine (äußere Membran: Porin-Moleküle; Poren-bildner, die die Membran für Moleküle durchlässig machen, die < 10 kDa sind)
 Anzahl und Gestalt der Mitochondrien sowie die Zahl der Cristae kann von Zelltyp zu Zelltyp unterschiedlich sein. Gewebe mit intensivem oxidativen Stoffwechsel besitzen Mitochondrien mit vielen Cristae-innerhalb eines Gewebes kann die Gestalt der Mitochondrien mit ihrem Funktionszustand variieren. Die Membranen der Mitochondrien enthalten integrale Proteine (äußere Membran: Porin-Moleküle; Poren-bildner, die die Membran für Moleküle durchlässig machen, die < 10 kDa sind)")
179
Energiebereitstellung
Karl-Franzens: Aufbau der Mitochondrien

180
Energiebereitstellung
EXKURS (Aufbau der Mitochondrien) Die innere Mitochondrienmembran ist für fast alle Moleküle undurchlässig (Ausnahmen: zB. O2, CO2, H2O) und weist einen Proteingehalt von ~ 75% auf: spezifische Carrier für den Stoffwechseltransport, Enzyme und andere Komponenten der Atmungskette und die ATP-Synthase Der Matrix-Raum ist reich an Proteinen, v.a. an Enzymen des Citrat-Zyklus
 Die innere Mitochondrienmembran ist für fast alle Moleküle undurchlässig (Ausnahmen: zB. O2, CO2, H2O) und weist einen Proteingehalt von ~ 75% auf: spezifische Carrier für den Stoffwechseltransport, Enzyme und andere Komponenten der Atmungskette und die ATP-Synthase. Der Matrix-Raum ist reich an Proteinen, v.a. an Enzymen des Citrat-Zyklus.")
181
Stoffwechselleistungen
Energiebereitstellung Stoffwechselleistungen Mitochondrien werden auch als die ‚biochemischen Kraftwerke‘ einer Zelle bezeichnet (sie erzeugen im Rahmen des oxidativen Abbaus von Nährstoffen den größten Teil des ATP‘s das die Zelle benötigt) Umwandlung von Pyruvat zu Acetyl-CoA Citrat-Zyklus Atmungskette (gekoppelt mit der Synthese von ATP = oxidative Phosphorylierung) Abbau der Fettsäuren durch -Oxidation Teile des Harnstoff-Zyklus Calciumspeicher Welche Prozesse finden in den Mitochondrien statt?
 Umwandlung von Pyruvat zu Acetyl-CoA. Citrat-Zyklus. Atmungskette (gekoppelt mit der Synthese von ATP = oxidative Phosphorylierung) Abbau der Fettsäuren durch -Oxidation. Teile des Harnstoff-Zyklus. Calciumspeicher. Welche Prozesse finden in den Mitochondrien statt")
182
Energiebereitstellung
Stoffwechselleistungen Die wesentliche Aufgabe der Mitochondrien: Aufnahme von Substraten des Energie-Stoffwechsels (FS, Pyruvat, C-Gerüste der AA‘s) aus dem Zytoplasma und oxidativer Abbau dieser Stoffe zu CO2 und H2O unter Gewinnung von ATP Die Reaktionen des Citratzyklus liefern vollständig oxidierten Kohlenstoff und Reduktionsäquivalente, die vorübergehend an Coenzyme gebunden sind (laufen vorwiegend im Matrixraum ab)
 aus dem Zytoplasma und oxidativer Abbau dieser Stoffe zu CO2 und H2O unter Gewinnung von ATP. Die Reaktionen des Citratzyklus liefern vollständig oxidierten Kohlenstoff und Reduktionsäquivalente, die vorübergehend an Coenzyme gebunden sind (laufen vorwiegend im Matrixraum ab)")
183
Energiebereitstellung
Stoffwechselleistungen Die Atmungskette benutzt NADH (Nicotinamid-Adenin-Dinucleotid) und das enzymgebundene FADH2 (Flavin-Adenin-Dinucleotid) als Reduktionsmittel (also Elektronenlieferanten) zur Reduktion von Sauerstoff und Bildung von Wasser. Diese Reaktion ist mit dem Transport von Protonen (H+) durch die innere Membran aus dem Matrix-Raum in den Intermembran-Raum gekoppelt
 und das enzymgebundene FADH2 (Flavin-Adenin-Dinucleotid) als Reduktionsmittel (also Elektronenlieferanten) zur Reduktion von Sauerstoff und Bildung von Wasser. Diese Reaktion ist mit dem Transport von Protonen (H+) durch die innere Membran aus dem Matrix-Raum in den Intermembran-Raum gekoppelt.")
184
Energiebereitstellung
Stoffwechselleistungen Bei diesem Vorgang wird ein elektrochemischer Gradient über die innere Membran aufgebaut – die Mitochondrien nutzen diesen Gradienten zur Synthese von ATP aus ADP u. Pi mit Hilfe der ATP-Synthase Exkurs: Elektrochemischer Gradient Elektrisch geladene Atome und Moleküle (Ionen) können biologische Membranen nur mit Hilfe von Ionenkanälen oder Ionenpumpen passieren. Die Ionenpumpen schaffen eine ungleiche Verteilung der Ionen auf beiden Seiten der Membran
 können. biologische Membranen nur mit Hilfe von Ionenkanälen oder Ionenpumpen passieren. Die Ionenpumpen schaffen eine ungleiche Verteilung der Ionen auf beiden Seiten der Membran.")
185
Energiebereitstellung
Stoffwechselleistungen Exkurs: Elektrochemischer Gradient Dadurch wird ein elektrochemischer Gradient aufgebaut (dieser speichert wie ein Kondensator Energie). Für die zelluläre ATP-Synthese sind Protonen-Gradienten wichtig. Der Energiegehalt eines solchen Gradienten hängt vom Unterschied der Protonenkonzentrationen auf beiden Seiten der Membran ab, d.h. von der pH-Differenz (pH) Ebenso leistet das Membranpotential (s)einen Beitrag. Aus beiden Größen zusammen ergibt sich die protonenmotorische Kraft (p) (Maß für die chemische Arbeit G die der Gradient leistet)
. Für die zelluläre ATP-Synthese sind Protonen-Gradienten wichtig. Der Energiegehalt eines solchen Gradienten hängt vom Unterschied der Protonenkonzentrationen auf beiden Seiten der Membran ab, d.h. von der pH-Differenz (pH) Ebenso leistet das Membranpotential (s)einen Beitrag. Aus beiden Größen zusammen ergibt sich die protonenmotorische Kraft (p) (Maß für die chemische Arbeit G die der Gradient leistet)")
187
Energiebereitstellung
Aerober Energiestoffwechsel Die Glycolyse ist ein kataboler Stoffwechselweg und findet im Zytoplasma statt: 1 mol Glucose wird dabei in 2 mol Pyruvat zerlegt. Zusätzlich entstehen bei diesem Vorgang je 2 mol ATP und NADH+ (wenn es sich aerobe Bedingungen handelt!) Unter anaeroben Bedingungen wird das Pyruvat weiter umgesetzt um das NAD+ zu regenerieren, dabei entsteht u.a. Laktat
 Unter anaeroben Bedingungen wird das Pyruvat weiter umgesetzt um das NAD+ zu regenerieren, dabei entsteht u.a. Laktat.")
188
Energiebereitstellung
Die Glucose wird in der Zelle zunächst phosphoryliert (Zucker werden vorwiegend als Phosphorsäure-Ester verstoffwechselt)
")
189
Energiebereitstellung
Die wesentlichen Abbauschritte Mit Hilfe der Hexokinase entsteht unter ATP-Verbrauch (1) G-6-P (Glukose-6-Phosphat) dieses wird isomerisiert zu (2) F-6-P (Fruktose-6-Phosphat); es wird erneut phosphorylisiert und es entsteht (3) F-1,6-BP (dieser Vorgang wird über das Enzym PFK (Phosphofruktokinase) katalysiert (wichtiges Schlüsselenzym der Glykolyse); bis zu diesem Punkt sind 2 mol ATP / mol Glucose verbraucht
 G-6-P (Glukose-6-Phosphat) dieses wird isomerisiert zu. (2) F-6-P (Fruktose-6-Phosphat); es wird erneut phosphorylisiert und es entsteht. (3) F-1,6-BP (dieser Vorgang wird über das Enzym PFK (Phosphofruktokinase) katalysiert (wichtiges Schlüsselenzym der Glykolyse); bis zu diesem Punkt sind 2 mol ATP / mol Glucose verbraucht.")
190
Energiebereitstellung
Die wesentlichen Abbauschritte F-1,6-BP wird nun durch die Aldolase in 2 phosphorylierte C3-Fragmente gespalten (4) Glyceral-3-phosphat und Glyceron-3-phosphat, diese werden (5) mittels der Triosephosphat-Isomerase ineinander übergeführt Glyc3-P wird nun durch die Glyceral-3-phosphat-Dehydrogenase unter Bildung von NADH+ + H+ oxidiert (6)
 Glyceral-3-phosphat und Glyceron-3-phosphat, diese werden. (5) mittels der Triosephosphat-Isomerase ineinander übergeführt. Glyc3-P wird nun durch die Glyceral-3-phosphat-Dehydrogenase unter Bildung von NADH+ + H+ oxidiert (6)")
191
Energiebereitstellung
Die wesentlichen Abbauschritte (6) Zusätzlich wird anorganisches Phosphat in das Molekül aufgenommen (‚Substratketten-phosphory-lierung‘) und es entsteht 1,3-Biphosphorglycerat (Intermediärprodukt) Im nächsten Schritt (7) wird die Hydrolyse der Bindung (gemischte Säureanhydridbindung) von 1,3-BPG (katalysiert durch die Phosphoglycerat-Kinase) mit der Bildung von ATP energetisch gekoppelt: 3-Phosphoglycerat
 Zusätzlich wird anorganisches Phosphat in das Molekül aufgenommen (‚Substratketten-phosphory-lierung‘) und es entsteht 1,3-Biphosphorglycerat (Intermediärprodukt) Im nächsten Schritt. (7) wird die Hydrolyse der Bindung (gemischte Säureanhydridbindung) von 1,3-BPG (katalysiert durch die Phosphoglycerat-Kinase) mit der Bildung von ATP energetisch gekoppelt: 3-Phosphoglycerat.")
192
Energiebereitstellung
Die wesentlichen Abbauschritte Ein weiteres Zwischenprodukt, dessen Hydrolyse sich mit der Synthese von ATP koppeln läßt, entsteht durch (8) Isomerisierung des 3-PG zu 2-Phosphoglycerat (Enzym: Phosphoglycerat-Mutase) und (9) H2O (Enzym: Phosphopyruvat-Hydratase, ‚Enolase‘). Das dabei gebildetet Produkt ist der Phosphorsäureester der Enol-Form des Pyruvats: Phosphoenolpyruvat
 Isomerisierung des 3-PG zu 2-Phosphoglycerat (Enzym: Phosphoglycerat-Mutase) und. (9) H2O (Enzym: Phosphopyruvat-Hydratase, ‚Enolase‘). Das dabei gebildetet Produkt ist der Phosphorsäureester der Enol-Form des Pyruvats: Phosphoenolpyruvat.")
193
Energiebereitstellung
Die wesentlichen Abbauschritte Im letzten Schritt dieses Prozesses (10) entsteht mittels der Pyruvat-Kinase aus PEP das Pyruvat Energetisch betrachtet: Nettogewinn des Abbaus: 2 (3) mol ATP / mol Glukose (d.h. ~ 31% der freien Energie werden für den ATP-Aufbau genutzt) 3 mol ATP entstehen, wenn Glykogen abgebaut wird
 entsteht mittels der Pyruvat-Kinase aus PEP das Pyruvat. Energetisch betrachtet: Nettogewinn des Abbaus: 2 (3) mol ATP / mol Glukose (d.h. ~ 31% der freien Energie werden für den ATP-Aufbau genutzt) 3 mol ATP entstehen, wenn Glykogen abgebaut wird.")
195
Energiebereitstellung
Die wesentlichen Abbauschritte Glykogen dient im tierischen Organismus als CHO-Reserve, aus der bei Bedarf Glucose-phosphate oder Glucose freigesetzt werden können Warum Glykogen als Speicherform? Die Speicherung von Glucose ist nicht sinnvoll, weil das Zellinnere stark hypertonisch wäre und es zu einem Einstrom von Wasser käme Da die osmotische Aktivität von der Zahl der Moleküle abhängt, sind Polysaccharide osmotisch fast inaktiv

196
Energiebereitstellung
Die wesentlichen Abbauschritte Der menschliche Organismus kann bis zu ~ 450g Glykogen speichern (1/3 davon in der Leber, der Rest in der Muskulatur) Wofür wird das Glykogen in der Leber benötigt? Das Leber-Glykogen dient primär der Aufrechterhaltung des Blutzuckerspiegels in der postresorptiven Phase und kann fast bis auf Null abfallen (beim Hungern!) Ab dann übernimmt die Glukoneogenese die Versorgung
 Wofür wird das Glykogen in der Leber benötigt Das Leber-Glykogen dient primär der Aufrechterhaltung des Blutzuckerspiegels in der postresorptiven Phase und kann fast bis auf Null abfallen (beim Hungern!) Ab dann übernimmt die Glukoneogenese die Versorgung.")
197
Energiebereitstellung
Die wesentlichen Abbauschritte Das Muskelglykogen dient hauptsächlich als Energiereserve (v.a. für die intensiven Belastungen im Kraftsport bzw. für den ‚Endspurt‘ bei den Ausdauersportarten) und ist nicht direkt an der Regulation des Blutzuckerspiegels beteiligt. Die Muskulatur besitzt keine G-6-phosphatase (damit kann die Glukose nicht ins Blut freigesetzt werden)
 und ist nicht direkt an der Regulation des Blutzuckerspiegels beteiligt. Die Muskulatur besitzt keine G-6-phosphatase (damit kann die Glukose nicht ins Blut freigesetzt werden)")
199
Energiebereitstellung
Die wesentlichen Abbauschritte Aufbau des Glykogen: Glykogen ist ein verzweigtes Homopolymer der Glukose (ähnelt baumartigen Strukturen, und kann bis aus Glukoseresten bestehen) Beim Abbau werden nur die nichtreduzierenden Enden des ‚Bäumchens‘ verkürzt / Wenn ein reichliches Glukoseangebot vorhanden ist, werden sie verlängert. Die Verlängerung wird durch das Enzym Glykogensynthase katalysiert
 Beim Abbau werden nur die nichtreduzierenden Enden des ‚Bäumchens‘ verkürzt / Wenn ein reichliches Glukoseangebot vorhanden ist, werden sie verlängert. Die Verlängerung wird durch das Enzym Glykogensynthase katalysiert.")
200
Energiebereitstellung
Die wesentlichen Abbauschritte Bei dieser Verlängerung wird ‚nur‘ bis zu einer bestimmten Länge der Kette ausgebaut und dann mittels eines verzweigenden Enzyms (Glucan verzweigendes Enzym) abgespalten und an einer benachbarten Kette angehängt Die verzweigte Struktur des Glykogens ermöglicht die rasche Freisetzung von Zucker-Resten. Das Enzym Glykogen-Phosphorylase spaltet vom nicht-reduzierenden Ende her nacheinander Reste als Glukose-1-phosphat ab.
 abgespalten und an einer benachbarten Kette angehängt. Die verzweigte Struktur des Glykogens ermöglicht die rasche Freisetzung von Zucker-Resten. Das Enzym Glykogen-Phosphorylase spaltet vom nicht-reduzierenden Ende her nacheinander Reste als Glukose-1-phosphat ab.")
201
Energiebereitstellung
Die wesentlichen Abbauschritte Je größer dabei die Zahl der Enden, desto mehr Phosphorylase-Moleküle können dabei gleichzeitig angreifen Die Bildung von G-1-P statt Glucose hat den Vorteil, daß kein ATP aufgewendet werden muß, um die freigesetzten Reste in die Glykolyse einzuschleusen

203
Energiebereitstellung
Die wesentlichen Abbauschritte Was passiert nun mit dem Pyruvat ? Das Pyruvat kann in den Matrix-Raum eindringen und dabei als Azetyl-Coenzym A in den Citratzyklus eingeschleust werden Das Azetyl-Coenzym A stammt aus der -Oxidation der Fettsäuren und aus der Pyruvat-Dehydrogenase Reaktion (Multienzymkomplex, der die oxidative Decarboxylierung von 2-Oxosäuren und die Übertragung des gebildeten Acyl-Restes auf Coenzym A katalysiert; als Elektronenakzeptor fungiert dabei NAD+)
")
204
Energiebereitstellung
Die wesentlichen Abbauschritte Citratzyklus Der Citratzyklus ist ein Stoffwechselweg der in der Mitochondrienmatrix abläuft; dieser Zyklus oxidiert Acetyl-Reste [CH3-CO-] zu CO2 Die gewonnenen Reduktionsäquivalente werden auf NAD+ oder Ubichinon übertragen und dort an die Atmungskette weitergegeben Die Oxidation des Acetyl-Restes verläuft über eine Reihe von Zwischenstufen:

205
Energiebereitstellung
Die wesentlichen Abbauschritte Citratzyklus (1) Der Acetyl-Rest wird auf das Trägermolekül Oxalacetat übertragen (katalysiert durch die Citrat-Synthase) [Vom Produkt dieser Reaktion, dem Citrat, hat der Zyklus seinen Namen] (2) Es erfolgt die Isomerisierung von Citrat zu Isocitrat und als Zwischenprodukt tritt Aconitat auf (katalysiert durch die Aconitat-Hydratase, ‚Aconitase‘)
 Der Acetyl-Rest wird auf das Trägermolekül Oxalacetat übertragen (katalysiert durch die Citrat-Synthase) [Vom Produkt dieser Reaktion, dem Citrat, hat der Zyklus seinen Namen] (2) Es erfolgt die Isomerisierung von Citrat zu Isocitrat und als Zwischenprodukt tritt Aconitat auf (katalysiert durch die Aconitat-Hydratase, ‚Aconitase‘)")
206
Energiebereitstellung
Die wesentlichen Abbauschritte Citratzyklus (3) Nun erfolgt der erste Oxidationsschritt - die Isocitrat-Dehydrogenase oxidiert die Hydroxy-Gruppe des Isocitrats zur Oxo-Gruppe. Glz. wird eine der Carboxy-Gruppen als CO2 abgespalten, und es entsteht 2-Oxoglutarat. (4) Der nächste Schritt ist die Umwandlung (umfasst eine Oxidation und eine Decarboxylierung) zum Succinyl-CoA und wird durch die 2-Oxoglutarat-Dehydrogenase katalysiert
 Nun erfolgt der erste Oxidationsschritt - die Isocitrat-Dehydrogenase oxidiert die Hydroxy-Gruppe des Isocitrats zur Oxo-Gruppe. Glz. wird eine der Carboxy-Gruppen als CO2 abgespalten, und es entsteht 2-Oxoglutarat. (4) Der nächste Schritt ist die Umwandlung (umfasst eine Oxidation und eine Decarboxylierung) zum Succinyl-CoA und wird durch die 2-Oxoglutarat-Dehydrogenase katalysiert.")
207
Energiebereitstellung
Die wesentlichen Abbauschritte Citratzyklus (5) Nachfolgend wird Succinyl-CoA in Succinat u. Coenzym A gespalten (katalysiert durch die Succinat-CoA-Ligase, ,Thiokinase‘). An diesem Punkt entsteht nicht ATP sondern Guanosin-triphosphat (GTP) das aber leicht in ATP umgewandelt werden kann. Bis hierher wird der Acetyl-Rest vollständig zu CO2 oxidiert. Gleichzeitig. wird das Trägermolekül Oxalacetat zu Succinat reduziert.
 Nachfolgend wird Succinyl-CoA in Succinat u. Coenzym A gespalten (katalysiert durch die Succinat-CoA-Ligase, ,Thiokinase‘). An diesem Punkt entsteht nicht ATP sondern Guanosin-triphosphat (GTP) das aber leicht in ATP umgewandelt werden kann. Bis hierher wird der Acetyl-Rest vollständig zu CO2 oxidiert. Gleichzeitig. wird das Trägermolekül Oxalacetat zu Succinat reduziert.")
208
Energiebereitstellung
Die wesentlichen Abbauschritte Citratzyklus In 3 weiteren Schritten des Zyklus wird nun aus Succinat wieder Oxalacetat regeneriert : (6) Zunächst wird durch die Succinat-Dehydrogenase Succinat zu Fumarat oxidiert. Die SDH ist ein integrales Protein der inneren Mitochondrienmembran (und muss der Atmungskette zugerechnet werden) Die SDH enthält FAD wobei bei dieser Reaktion der eigentliche Elektronen-Akzeptor das Ubichinon ist
 Zunächst wird durch die Succinat-Dehydrogenase Succinat zu Fumarat oxidiert. Die SDH ist ein integrales Protein der inneren Mitochondrienmembran (und muss der Atmungskette zugerechnet werden) Die SDH enthält FAD wobei bei dieser Reaktion der eigentliche Elektronen-Akzeptor das Ubichinon ist.")
209
Energiebereitstellung
Die wesentlichen Abbauschritte Citratzyklus (7) An die Doppelbindung des Fumarats wird nun durch die Fumarat-Hydratase (‚Fumarase‘) Wasser addiert und es ensteht Malat Im letzten Schritt des Zyklus wird Malat durch die Malat-Dehydrogenase wieder zu Oxalacetat oxidiert, wobei noch einmal NADH+ + H+ entsteht ….somit ist der Kreis geschlossen, und der Zyklus kann von vorne beginnen…..
 An die Doppelbindung des Fumarats wird nun durch die Fumarat-Hydratase (‚Fumarase‘) Wasser addiert und es ensteht Malat. Im letzten Schritt des Zyklus wird Malat durch die Malat-Dehydrogenase wieder zu Oxalacetat oxidiert, wobei noch einmal NADH+ + H+ entsteht. ….somit ist der Kreis geschlossen, und der Zyklus. kann von vorne beginnen…..")
210
Citratzyklus

211
Energiebereitstellung
Karl-Franzens: Zur Erinnerung…. Die ATP-Produktion muß ständig dem wechselnden Bedarf angepasst werden Der Wiederaufbau des ATP aus ADP erfolgt auf 3 Wegen Aus KP = anaerob alaktazide Resynthese Über die anaerobe Oxidation = anaerob laktazide Resynthese Über die aerobe Oxidation = aerobe Resynthese

212
Energiebereitstellung
Karl-Franzens: Der kleinere Energiespeicher (ATP) beliefert DIREKT die energiebenötigenden Reaktionen Der größere Energiespeicher (KP) füllt den ATP-Speicher wieder auf Bei einem ausreichenden Sauerstoffangebot (aerobe Oxidation) erfolgt der Abbau von KH und Fetten (bzw. Proteinen) SCHRITTWEISE in enzymgesteuerten Stufen zu CO2 und H2O (energie- freie Endprodukte)
 beliefert DIREKT die energiebenötigenden Reaktionen. Der größere Energiespeicher (KP) füllt den ATP-Speicher wieder auf. Bei einem ausreichenden Sauerstoffangebot. (aerobe Oxidation) erfolgt der Abbau von. KH und Fetten (bzw. Proteinen) SCHRITTWEISE in. enzymgesteuerten Stufen zu CO2 und H2O (energie- freie Endprodukte)")
213
Energiebereitstellung
Karl-Franzens: Aerobe Oxidation: Glykogenolyse (Glykogenabbau) Glykolyse (Glukoseabbau) Bildung der aktivierten Essigsäure (Azetyl-CoA) damit das gebildete Pyruvat für den weiteren Abbau verwendet werden kann Trikarbonsäure- oder Citrazyklus (auch Krebszyklus genannt) Atmungskette (oder Elektronentransportkette)
 Glykolyse (Glukoseabbau) Bildung der aktivierten Essigsäure (Azetyl-CoA) damit das gebildete Pyruvat für den weiteren Abbau verwendet werden kann. Trikarbonsäure- oder Citrazyklus (auch Krebszyklus genannt) Atmungskette (oder Elektronentransportkette)")
214
Energiebereitstellung
Karl-Franzens: Wenn das Sauerstoffangebot nicht ausreicht, dann wird der Weg der anaeroben Glykolyse beschritten [dann, wenn der momentane Energiebedarf nicht durch aerobe Oxidation gedeckt werden kann] Die maximale aerobe Oxidation wird begrenzt: O2-Menge, die der Muskelzelle/Zeiteinheit zugeführt werden kann Menge der Enzyme, die die einzelnen Reaktionen bei der Oxidation katalysieren Größe der Nährstoffdepots in der Muskelzelle sowie deren Wiederauffüllung

215
Energiebereitstellung
Anaerobe Energiebereitstellung In Abwesenheit von Sauerstoff (zeitabhängige Transportprozeße an der inneren Mitochondrien-membran – z.T. TRAINIERBAR ! – durch Zunahme der Oberfläche und tlw. genetisch bestimmt aufgrund der Muskelfaserverteilung) kann in den Mitochondrien NADH nicht mehr reoxidiert werden; damit kommt außer der ATP-Bildung auch der Citrat-Zyklus (und damit der FS-und AS-Abbau) zum Erliegen Ort des Geschehens: die Enzyme befinden sich im Sarkoplasma (in unmittelbarer Nähe der Myofibrillen)
 kann in den Mitochondrien NADH nicht mehr reoxidiert werden; damit kommt außer der ATP-Bildung auch der Citrat-Zyklus (und damit der FS-und AS-Abbau) zum Erliegen. Ort des Geschehens: die Enzyme befinden sich im Sarkoplasma (in unmittelbarer Nähe der Myofibrillen)")
216
Energiebereitstellung
Anaerobe Energiebereitstellung Als Energiesubstrat bleibt dann nur mehr Glukose übrig [der Abbau im Rahmen der Glykolyse zu Pyruvat (ionisierte Form der Brenztraubensäure) liefert 2 mol ATP] Wenn dieser Prozeß kontinuierlich weiter laufen soll (wenn also bei hoher Intensität die Leistung aufrecht erhalten werden muß), dann muß das in der Glykolyse gebildete NADH ausserhalb der Mitochondrien regeneriert werden – dazu wird Pyruvat zu Laktat reduziert
 liefert 2 mol ATP] Wenn dieser Prozeß kontinuierlich weiter laufen soll (wenn also bei hoher Intensität die Leistung aufrecht erhalten werden muß), dann muß das in der Glykolyse gebildete NADH ausserhalb der Mitochondrien regeneriert werden – dazu wird Pyruvat zu Laktat reduziert.")
217
Energiebereitstellung
Anaerobe Energiebereitstellung Warum ist das so? Wird die Glykolyse durch eine starke Steigerung des Energiebedarfs gefordert, so fällt mehr Brenztraubensäure an, als oxidativ verarbeitet werden kann; dadurch wird die Glykolyse verlangsamt und damit die anaerobe Energiebereitstellung gefährdet [Ursache: neben Brenztraubensäure häuft sich NADH+H+ (reduzierte Form) an – dieses muss aber als NAD+ vorliegen (in oxidierter Form) um als H+-Akzeptor fungieren zu können]
 an – dieses muss aber als NAD+ vorliegen (in oxidierter Form) um als H+-Akzeptor fungieren zu können]")
218
Energiebereitstellung
Anaerobe Energiebereitstellung Ausweg: Da die Atmungskette nicht so rasch das anfallende NADH+H+ oxidieren kann wird Wasserstoff des NADH+H+ auf Pyruvat übertragen, wobei Laktat und das benötigte NAD+ entsteht De facto ist die AN-Glykolyse ein Stoffwechselweg, der unabhängig vom aktuellen Zustand der aeroben Oxidation ablaufen kann; Probleme macht die Tatsache, dass die [H+] proportional zur Laktatkonzentration ansteigt (unter physiolog. Bedingungen ist Laktat zu über 95% dissoziiert)
")
219
Karl-Franzens:

220
Energiebereitstellung
Anaerobe Energiebereitstellung Das Enzym das die Reduktion zu Laktat katalysiert ist die Laktatdehydrogenase (2 verschiedene LDH Untereinheiten: H- bzw. M-Form, und 5 Isoenzyme der LDH) In der Herz-Muskulatur findet sich vorwiegend Tetramere der H-Form, in der LDH der Leber und der Skelettmuskulatur vorwiegend M-Monomere
 In der Herz-Muskulatur findet sich vorwiegend Tetramere der H-Form, in der LDH der Leber und der Skelettmuskulatur vorwiegend M-Monomere.")
221
Energiebereitstellung
Anaerobe Energiebereitstellung Die LDH katalysiert die Übertragung von Redox-Äquivalenten von Laktat auf NAD+ oder von NADH auf Pyruvat Laktat + NAD+ Pyruvat + NADH + H+ Die LDH katalysiert die Reaktion in beide Richtungen, hat aber keinen Einfluß auf die Lage des chem. Gleichgewichts; das Gleichgewicht der Reaktion liegt aber stark auf der Seite der Laktat-Bildung; bei hohen Konzentrationen von Laktat und NAD+ ist aber auch die Oxidation von Laktat zu Pyruvat möglich

222
Energiebereitstellung
Karl-Franzens: Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Das wichtigste regulierende Enzym der Glykolyse ist die PFK; sie wird durch ADP, AMP, und F-6-P als Substrat aktiviert und durch ATP und H+-Ionen gehemmt. Sinkt der pH-Wert auf z.B. 6.7 ab, so beträgt die Aktivität der PFK (und damit die Flußrate) nur mehr ~ 50% des Maximums; bei einem pH-Wert von ~ 6.3 ist sie komplett gehemmt (die ATP-Resyntheserate aus der Glykolyse geht gegen Null)
 nur mehr ~ 50% des Maximums; bei einem pH-Wert von ~ 6.3 ist sie komplett gehemmt (die ATP-Resyntheserate aus der Glykolyse geht gegen Null)")
223
Energiebereitstellung
Karl-Franzens: Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Die pH-bedingte Hemmung der Glykolyse schützt vor der zunehmenden Aktivierung von Lysosomen [ µm große Organellen, die dem Abbau von Zellbestandteilen dienen] – damit wird eine „Selbstverdauung“ der Zelle verhindert Die Übersäuerung der Zelle kann verzögert werden durch: 1. die in der Zelle vorhandenen Puffersysteme 2. Die Entfernung des Laktats

224
Energiebereitstellung
Karl-Franzens: Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Das anfallende Laktat wird bereits während der Arbeit weiterverwertet (v.a. bei ausreichend durchbluteter Muskulatur); in der Arbeitsmuskulatur (hptsl. in den roten MF und in den weniger stark belasteten Faseranteilen, die sich außerhalb der Belastungsachsen befinden), in der nicht-arbeitenden Muskulatur, in der Herzmuskulatur (der Herzmuskel ist aufgrund seiner aeroben Leistungsfähigkeit in der Lage, einen großen Prozentsatz seines Energiebedarfs über Laktat abzudecken)
; in der Arbeitsmuskulatur (hptsl. in den roten MF und in den weniger stark belasteten Faseranteilen, die sich außerhalb der Belastungsachsen befinden), in der nicht-arbeitenden Muskulatur, in der Herzmuskulatur (der Herzmuskel ist aufgrund seiner aeroben Leistungsfähigkeit in der Lage, einen großen Prozentsatz seines Energiebedarfs über Laktat abzudecken)")
225
Energiebereitstellung
Karl-Franzens: Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Ein Teil des Laktats wird in der Leber wieder zu Glukose aufgebaut (Glukoneogenese) [Dabei wird das im Blut zirkulierende Laktat zur Leber transportiert und im Rahmen des CORI-Zyklus zu Glukose umgewandelt und wieder zum Verbraucher zurücktransportiert]
 [Dabei wird das im Blut zirkulierende Laktat zur Leber transportiert und im Rahmen des CORI-Zyklus zu Glukose umgewandelt und wieder zum Verbraucher zurücktransportiert]")
226
Energiebereitstellung
Karl-Franzens: Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Der messbare Laktatanstieg im Blut wäre noch größer, wenn nicht ein Teil des anfallenden Pyruvats durch Aminierung (Substitutionsreaktion) in die AS Alanin übergeführt würde (Alanin wird in der Leber - aber nicht im Muskel - zu Glukose und Glykogen aufgebaut) – Alanin-Zyklus (In der Erholungsphase kann das durch anerobe Belastung gebildete Laktat im Muskel zu ~ 50% über die Glukoneogenese wieder zu Glukose und Glykogen umgewandelt werden)
 in die AS Alanin übergeführt würde (Alanin wird in der Leber - aber nicht im Muskel - zu Glukose und Glykogen aufgebaut) – Alanin-Zyklus. (In der Erholungsphase kann das durch anerobe Belastung gebildete Laktat im Muskel zu ~ 50% über die Glukoneogenese wieder zu Glukose und Glykogen umgewandelt werden)")
227
Energiebereitstellung
Karl-Franzens: Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Der Alanin-Zyklus wird durch den Abbau von Proteinen gespeist - die dabei anfallenden AS werden durch Transaminierung in 2-Oxosäuren umgewandelt, die großteils in den Citratzyklus wandern; dabei werden ihre Amino-Gruppen auf Pyruvat übertragen, das in Alanin übergeht (der Alanin-Zyklus dient glz. dem Transport von Glukose-Vorstufen und Stickstoff zur Leber – in der Leber wird N zu Harnstoff umgesetzt)
")
228
Karl-Franzens:

229
Energiebereitstellung
Karl-Franzens: Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Nach intensiven Belastungen (z.B. 400m-Lauf) steigt das Blutlaktat noch weiter an, und erreicht in Abhängigkeit von Belastungsdauer und [LA] ein Maximum nach 1-10min. Erholungszeit – die Ursache dafür ist der Konzentrationsgradient zwischen Muskel- und Blutlaktatkonzentration Für die Erfassung der LA-Elimination wird die Halbwertszeit (Zeit, nach der das LA auf die Hälfte des max. Wertes abgebaut ist) angegeben (diese hängt von der Höhe des maximal erreichten LA-Wertes ab!)
 steigt das Blutlaktat noch weiter an, und erreicht in Abhängigkeit von Belastungsdauer und [LA] ein Maximum nach 1-10min. Erholungszeit – die Ursache dafür ist der Konzentrationsgradient zwischen Muskel- und Blutlaktatkonzentration. Für die Erfassung der LA-Elimination wird die Halbwertszeit (Zeit, nach der das LA auf die Hälfte des max. Wertes abgebaut ist) angegeben (diese hängt von der Höhe des maximal erreichten LA-Wertes ab!)")
230
Laktatbildung und LA Verhalten im Muskel und im Blut
Karl-Franzens: Laktatbildung und LA Verhalten im Muskel und im Blut (de Marées, H.: Sportphysiologie)
")
231
Energiebereitstellung
Karl-Franzens: Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Im Mittel geht man von ½-T von 15min. bei einer [LA] von 10mmol aus; bei sehr hohen [LA] kann ½-T 25min. überschreiten Die Ursache für diese konzentrationsabhängige ½-T wird in der Existenz eines ‚Laktat-Shuttle‘ Systems gesehen; dabei dürfte das LA mittels spezifischer Proteine durch Zellmembranen transportiert werden; die Transportrate ist dabei eine nichtlineare Funktion der [LA]

232
Laktatverhalten im Blut (1/2-T) bei unterschiedlichen
Karl-Franzens: Laktatverhalten im Blut (1/2-T) bei unterschiedlichen LA-Ausgangswerten (de Marées, H.: Sportphysiologie)
 bei unterschiedlichen. LA-Ausgangswerten (de Marées, H.: Sportphysiologie)")
233
Energiebereitstellung
Karl-Franzens: Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Aktive Erholung (leichte Belastung) verkürzt die ½-T der LA-Elimination! Maximale Eliminationsraten erreicht man bei Belastungen von 40-50% der maximalen VO2 [durch den erhöhten Muskelstoffwechsel wird Laktat vermehrt in Brenztraubensäure übergeführt und in weiterer Folge als direkt energielieferndes Substrat mittels aerober Oxidation verstoffwechselt; d.h. Laktat ist energetisch ‚wertvoll‘
 verkürzt die ½-T der LA-Elimination! Maximale Eliminationsraten erreicht man bei Belastungen von 40-50% der maximalen VO2 [durch den erhöhten Muskelstoffwechsel wird Laktat vermehrt in Brenztraubensäure übergeführt und in weiterer Folge als direkt energielieferndes Substrat mittels aerober Oxidation verstoffwechselt; d.h. Laktat ist energetisch ‚wertvoll‘")
234
Karl-Franzens: Einfluß von passiver Erholung und aktiver Erholung auf das Blutlaktat nach einer Belastung (de Marées, H.: Sportphysiologie)
")
235
Energiebereitstellung
Karl-Franzens: Anaerobe Energiebereitstellung Was passiert mit dem Laktat? Laktat wird im Stoffwechsel immer gebildet (Ruhelaktatwerte von ~ mmol/l), und diese Bildung hängt u.a. auch von der Ernährung ab Die LA-Elimination hängt von der Höhe des aeroben Umsatzes und von der [LA] ab. Je höher der aerobe Stoffwechsel und je höher die [LA], desto größer die Elimination Die jeweils aktuelle [LA] im Muskel, im Blut und in anderen Körperflüssigkeiten ist das Resultat von LA-bildung,-diffusion,-transport, und -elimination
, und diese Bildung hängt u.a. auch von der Ernährung ab. Die LA-Elimination hängt von der Höhe des aeroben Umsatzes und von der [LA] ab. Je höher der aerobe Stoffwechsel und je höher die [LA], desto größer die Elimination. Die jeweils aktuelle [LA] im Muskel, im Blut und in. anderen Körperflüssigkeiten ist das Resultat von. LA-bildung,-diffusion,-transport, und -elimination.")
236
Energiebereitstellung
Karl-Franzens: Anaerobe Energiebereitstellung Im Vergleich zum Gesamtgewinn von 31 mol ATP (wenn 1 mol Glukose abgebaut wird) bei der aeroben Oxidation, ergeben sich für die anaerobe Glykolyse 3 mol ATP/mol Glukose (~ 10%) Da die maximale Umsatzrate der Glukose jedoch 25fach höher ist als die von Citratzyklus und Atmungskette (hier geht man von den Werten für Untrainierte aus), resultiert eine etwa doppelt so große ATP-Resynthese/Zeiteinheit aus der anaeroben Glykolyse
 bei der aeroben Oxidation, ergeben sich für die anaerobe Glykolyse 3 mol ATP/mol Glukose (~ 10%) Da die maximale Umsatzrate der Glukose jedoch 25fach höher ist als die von Citratzyklus und Atmungskette (hier geht man von den Werten für Untrainierte aus), resultiert eine etwa doppelt so große ATP-Resynthese/Zeiteinheit aus der anaeroben Glykolyse.")
237
Vorteile/Nachteile der aeroben und anaeroben Energiebereitstellung
Karl-Franzens: Vorteile/Nachteile der aeroben und anaeroben Energiebereitstellung Aerobe EB Anaerobe EB 31 mol ATP/mol Glukose 3 mol ATP/mol Glukose Keine LA-Bildung LA-Bildung (-) langsame Energie-bereitstellung (+) schnelle Energie-bereitstellung (-) freigesetzte Energ.-menge/Zeiteinheit ist relativ klein (+) freigesetzte Energ.menge/Zeiteinheit ist relativ groß
 langsame Energie-bereitstellung. (+) schnelle Energie-bereitstellung. (-) freigesetzte Energ.-menge/Zeiteinheit ist relativ klein. (+) freigesetzte Energ.menge/Zeiteinheit ist relativ groß.")
238
Vorteile/Nachteile der aeroben und anaeroben Energiebereitstellung
Karl-Franzens: Vorteile/Nachteile der aeroben und anaeroben Energiebereitstellung Aerobe EB Anaerobe EB (+) bereitgestellte Energiemenge ist relativ groß (-) die Gesamtenergiemenge ist relativ klein Sportliche Tätigkeiten können relativ lange durchgehalten werden, wenn die Intensität nicht zu hoch ist (1000m Lauf) Sportliche Tätigkeiten können mit hoher Intensität geleistet werden, aber nur kurzzeitig (400m Lauf)
 bereitgestellte Energiemenge ist relativ groß. (-) die Gesamtenergiemenge ist relativ klein. Sportliche Tätigkeiten können relativ lange durchgehalten werden, wenn die Intensität nicht zu hoch ist (1000m Lauf) Sportliche Tätigkeiten können mit hoher Intensität geleistet werden, aber nur kurzzeitig (400m Lauf)")
239
(Markworth, P. SPORTMEDIZIN)
Karl-Franzens: Laktatverhalten und Anteile der Energie-gewinnung bei unterschiedlichen (Markworth, P. SPORTMEDIZIN) Belastungsintensitäten
 Belastungsintensitäten.")
240
Karl-Franzens: Anteil der energiebereitstellenden Systeme im zeitl.Verlauf einer körperlichen Belastung (Markworth, P. SPORTMEDIZIN)
 .")
241
ENDOKRINOLOGIE Womit beschäftigt sich die Endokrinologie? Die Endokrinologie ist die Lehre von den Hormonen. Der klassischen Definition nach sind Hormone chemische Nachrichtenträger, die in spezialisierten Zellen (Drüsenzellen) produziert werden. Von dort werden sie in das Blut sezerniert und gelangen zu ihren Zielorganen oder Zielzellen, wo sie ihre spezifische Wirkung(en) vermitteln. Diese Zielorgane (Zielzellen) besitzen entsprechende Rezeptoren für das entsprechende Hormon
 produziert werden. Von dort werden sie in das Blut sezerniert und gelangen zu ihren Zielorganen oder Zielzellen, wo sie ihre spezifische Wirkung(en) vermitteln. Diese Zielorgane (Zielzellen) besitzen entsprechende Rezeptoren für das entsprechende Hormon.")
242
Hormonbildende Drüsen

243
a.) kurzfristig (innerhalb von Minuten)
ENDOKRINOLOGIE Was machen bzw. bewirken Hormone? Hormone beeinflussen im Körper verschiedene Funktionen; die Initialisierung und Regelung dieser Funktionen erfolgt dabei a.) kurzfristig (innerhalb von Minuten) b.) mittelfristig (innerhalb von Stunden) c.) langfristig (innerhalb von Tagen) Die hormonelle Nachrichtenübermittlung dauert daher wesentlich länger als die nervale
 kurzfristig (innerhalb von Minuten) b.) mittelfristig (innerhalb von Stunden) c.) langfristig (innerhalb von Tagen) Die hormonelle Nachrichtenübermittlung dauert daher wesentlich länger als die nervale.")
244
ENDOKRINOLOGIE Klassifizierung von Hormonen Hormone sind entweder Proteine bzw. Aminosäurenderivate, oder Lipide – entsprechend nach der Rezeptorlokalisation in den Zielzellen lassen sich 3 Arten einteilen: (1) Protein-, Proteo-, und Peptidhormone (hydrophil) bestehen aus AS und können die Lipidbarriere der Plasmazellmembran kaum passieren. Die Rezeptoren für diese Hormone sitzen deshalb an der Oberfläche der Plasmazellmembran
 Protein-, Proteo-, und Peptidhormone (hydrophil) bestehen aus AS und können die Lipidbarriere der Plasmazellmembran kaum passieren. Die Rezeptoren für diese Hormone sitzen deshalb an der Oberfläche der Plasmazellmembran.")
245
ENDOKRINOLOGIE Klassifizierung von Hormonen (2) Lipidhormone (lipophil) können aufgrund ihrer guten Löslichkeit durch die Zellmembran diffundieren und binden an Rezeptoren (diese sind meistens im Zytoplasma lokalisiert) (3) Schilddrüsenhormone entstehen aus 2 AS die über eine Etherbrücke verbunden sind; SH können gut in alle Körperzellen eindringen und binden an Rezeptoren im Zellkern Eine Zelle kann glz. Rezeptoren aller drei Typen besitzen bzw. auch über unterschiedliche Rezeptoren eines Typs verfügen
 Lipidhormone (lipophil) können aufgrund ihrer guten Löslichkeit durch die Zellmembran diffundieren und binden an Rezeptoren (diese sind meistens im Zytoplasma lokalisiert) (3) Schilddrüsenhormone entstehen aus 2 AS die über eine Etherbrücke verbunden sind; SH können gut in alle Körperzellen eindringen und binden an Rezeptoren im Zellkern. Eine Zelle kann glz. Rezeptoren aller drei Typen besitzen bzw. auch über unterschiedliche Rezeptoren eines Typs verfügen.")
246
ENDOKRINOLOGIE Wirkmechanismen von Hormonen Prinzipiell werden 3 Formen bzw. Wirkungen der Signalübertragung unterschieden: 1. Endokrine Wirkung: das Hormon überträgt das Signal durch Wanderung (über das Blut) vom Ort seiner Bildung zum Ort seiner Wirkung („klassische hormonelle Regulation“; z.B. INSULIN)
 vom Ort seiner Bildung zum Ort seiner Wirkung („klassische hormonelle Regulation ; z.B. INSULIN)")
247
ENDOKRINOLOGIE Wirkmechanismen von Hormonen 2. Parakrine Wirkung: das Hormon wirkt auf Zielzellen, die sich in unmittelbarer Nähe der Drüsenzelle befindet (z.B. lokal wirkende Gewebshormone, wie des Gastrointestinaltraktes) Autokrine Wirkung: das Hormon wirkt auf die Drüsenzelle selbst (Autoregulation) Ein (auch klassisches) Hormon kann sowohl endokrin, parakrin als auch autokrin wirken
 Autokrine Wirkung: das Hormon wirkt auf die Drüsenzelle selbst (Autoregulation) Ein (auch klassisches) Hormon kann sowohl endokrin, parakrin als auch autokrin wirken.")
248
Wirkmechanismen von Hormonen

249
ENDOKRINOLOGIE Wirkmechanismen von Hormonen Aufgrund der Interaktion von Hormon und Rezeptor (entweder an der Zellmembran oder in der Zelle) gibt es 2 völlig unterschiedliche Wirkmechanismen der Hormone: Intrazelluläre Hormon-Rezeptor-Komplex übt fast immer eine direkt genomische Wirkung aus. Diese genomischen Effekte (‚Transkriptionskontrolle‘) beeinflussen wiederum die Syntheseleistung von mRNA der Zelle [Steroidhormone, Thyroxin]
 gibt es 2 völlig unterschiedliche Wirkmechanismen der Hormone: Intrazelluläre Hormon-Rezeptor-Komplex übt fast immer eine direkt genomische Wirkung aus. Diese genomischen Effekte (‚Transkriptionskontrolle‘) beeinflussen wiederum die Syntheseleistung von mRNA der Zelle [Steroidhormone, Thyroxin]")
250
ENDOKRINOLOGIE Wirkmechanismen von Hormonen 2. Plasmazellmembranständige Hormon-Rezeptor-Komplex wirkt über second messengers (cAMP, cGMP, DAG, IP3, Ca2+, Arachidonat). Dabei wird über die spezifische Botschaft des Hormons und spezifische Rezeptoren ein unspezifischer intrazellulärer Mechanismus ausgelöst. Über weitere Reaktionen werden dann die Antworten der Zielzelle auf das Hormon gesteuert
. Dabei wird über die spezifische Botschaft des Hormons und spezifische Rezeptoren ein unspezifischer intrazellulärer Mechanismus ausgelöst. Über weitere Reaktionen werden dann die Antworten der Zielzelle auf das Hormon gesteuert.")
251
Die Bindung von Hormonen an Rezeptoren in der
ENDOKRINOLOGIE Wirkmechanismen von Hormonen Die Bindung von Hormonen an Rezeptoren in der Zellwand, im Zellplasma oder im Zellkern (und damit auch verbunden die Wirksamkeit) ist abhängig von der Rezeptorspezifität der Rezeptoraffinität (Bindungsstärke) und der Rezeptordichte Die R.Dichte variiert in Abhängigkeit von der jeweiligen Hormon-Konzentration: Sind die Hormonkonzentrationen über längere Zeit erhöht, erfolgt eine DOWN-REGULATION der aktiven Rezeptoren (umgekehrt: UP-REGULATION)
 ist. abhängig von. der Rezeptorspezifität. der Rezeptoraffinität (Bindungsstärke) und. der Rezeptordichte. Die R.Dichte variiert in Abhängigkeit von der jeweiligen Hormon-Konzentration: Sind die Hormonkonzentrationen über längere Zeit erhöht, erfolgt eine DOWN-REGULATION der aktiven Rezeptoren (umgekehrt: UP-REGULATION)")
252
Hierarchische Struktur
und Aufbau der Hypothalamus- Hypophysen-Hormondrüsen Achse

253
Hypothalamus-Hypophysen System
ENDOKRINOLOGIE Hypothalamus-Hypophysen System Der Hypothalamus ist eine zentrale Region für die Steuerung der vegetativen Funktionen und beeinflusst auch das endokrin-vaskuläre System („nerval-hormonelle Schaltstelle“) Zusammen mit der Hypophyse [v.a. mit der Adenohypophyse (Hypophysenvorderlappen)] fungiert der Hypothalamus als übergeordnete Einheit eines mehrstufigen, hierarchisch-strukturierten Systems (! eine Reihe von Hormonwirkungen können aber unabhängig vom H-H System ablaufen !)
 Zusammen mit der Hypophyse [v.a. mit der Adenohypophyse (Hypophysenvorderlappen)] fungiert der Hypothalamus als übergeordnete Einheit eines mehrstufigen, hierarchisch-strukturierten Systems (! eine Reihe von Hormonwirkungen können aber unabhängig vom H-H System ablaufen !)")
255
Funktionelle Unterteilung der Hormone
ENDOKRINOLOGIE Hypothalamus-Hypophysen System Funktionelle Unterteilung der Hormone die im mittleren Teil des Hypothalamus gebildeten Hormone haben die Aufgabe, in der Adenohypophyse die Sekretion (bzw. auch die Bildung) weiterer Hormone anzuregen (Releasinghormone, RH) oder zu hemmen (Inhibiting-Hormone, IH) einige der in der Adenohypophyse gebildeten Hormone (glandotropen H) wirken auf die peripheren Hormondrüsen
 weiterer Hormone anzuregen (Releasinghormone, RH) oder zu hemmen (Inhibiting-Hormone, IH) einige der in der Adenohypophyse gebildeten Hormone (glandotropen H) wirken auf die peripheren Hormondrüsen.")
256
ENDOKRINOLOGIE Hypothalamus-Hypophysen System die peripheren Hormondrüsen schütten am Ende dieser Kaskade die glandulären Hormone aus, die dann letztendlich an den Zielzellen unterschiedlicher Organe wirksam werden Jene Hormone, die direkt am Erfolgsorgan wirken, werden auch als effektorische Hormone (oder Effektorhormone) bezeichnet
 bezeichnet.")
257
Rückkoppelungssysteme-Hormonelle Regelkreise
ENDOKRINOLOGIE Wirkmechanismen von Hormonen Rückkoppelungssysteme-Hormonelle Regelkreise Endokrine Regulationen lassen sich durch einen Regelkreis beschreiben (der ‚Erfolg‘ der Funktion eines Hormons wird dem regelnden Teil unmittelbar oder mittelbar zurückgemeldet) Negative Rückkoppelung: die Konzentration eines vorhandenen H im Blut wird ‚gemessen‘, und dieser Wert wird an das oberste Regelzentrum (Hypothalamus) und die Adenohypophyse ‚gemeldet‘ = IST-Wert
 Negative Rückkoppelung: die Konzentration eines vorhandenen H im Blut wird ‚gemessen‘, und dieser Wert wird an das oberste Regelzentrum (Hypothalamus) und die Adenohypophyse ‚gemeldet‘ = IST-Wert.")
258
ENDOKRINOLOGIE Wirkmechanismen von Hormonen Negative Rückkoppelung Im Hypothalamus wird ein IST-SOLLWert Vergleich vorgenommen; dieser führt zu einer Verringerung der RH-Freisetzung (od. zu einer Erhöhung der IH-Freisetzung) worauf die Produktion/Freisetzung des betreffenden glandotropen H (= REGELGRÖSSE) verringert wird
 worauf die Produktion/Freisetzung des betreffenden glandotropen H (= REGELGRÖSSE) verringert wird.")
259
ENDOKRINOLOGIE Wirkmechanismen von Hormonen Positive Rückkoppelung (sind selten): die Zunahme eines H (z.B. FSH) führt zu einer verstärkten Produktion eines effektorischen H (z.B. Östradiol) das wiederum die FSH – (und LH) Ausschüttung stimuliert (Ovulation) Oxytozin fördert die Kontraktion der Uterusmuskulatur; dies führt auf nervalem Wege zu einer Rückmeldung an den Hypophysenhinterlappen: Verstärkung der Oxytozinausschüttung
: die Zunahme eines H (z.B. FSH) führt zu einer verstärkten Produktion eines effektorischen H (z.B. Östradiol) das wiederum die FSH – (und LH) Ausschüttung stimuliert (Ovulation) Oxytozin fördert die Kontraktion der Uterusmuskulatur; dies führt auf nervalem Wege zu einer Rückmeldung an den Hypophysenhinterlappen: Verstärkung der Oxytozinausschüttung.")
260
Rückkopplungsprinzip
von Hormonen

261
Hormone des Hypophysenvorderlappens
ENDOKRINOLOGIE Hormone des Hypophysenvorderlappens Glandotrope H: TSH Thyreotropin (Schilddrüse), ACTH Kortikotropin (Nebennierenrinde), FSH Follitropin (Keimdrüsen), LH Lutropin (Keimdrüsen) [LH, FSH Gonadotropine]; für jedes dieser H gibt es ein RH Effektor H: STH Somatotropin (wirkt auf viele Gewebe (Muskel, Fett, Knochen, Knorpel), PRL Prolaktin (IH: Prolaktostatin = Dopamin), -MSH Melanotropin, -Endorphin
, ACTH Kortikotropin (Nebennierenrinde), FSH Follitropin (Keimdrüsen), LH Lutropin (Keimdrüsen) [LH, FSH Gonadotropine]; für jedes dieser H gibt es ein RH. Effektor H: STH Somatotropin (wirkt auf viele Gewebe (Muskel, Fett, Knochen, Knorpel), PRL Prolaktin (IH: Prolaktostatin = Dopamin), -MSH Melanotropin, -Endorphin.")
262
Hormone des Hypophysenhinterlappens
ENDOKRINOLOGIE Hormone des Hypophysenhinterlappens Effektor H: ADH Adiuretin (wirkt in den Nieren-sorgt u.a. dafür, dass dem Körper möglichst wenig an Wasser verloren geht), Oxytozin Periphere Hormondrüsen SCHILDDRÜSE ist der Bildungs- und Aus-schüttungsort der Hormone Thyroxin, Trijodthyronin (Steigerung des Energieumsatzes, Förderung der Glykogenspaltung, Fettmobilisierung), Kalzitonin (Kalziumhaushalt)
, Oxytozin. Periphere Hormondrüsen. SCHILDDRÜSE ist der Bildungs- und Aus-schüttungsort der Hormone Thyroxin, Trijodthyronin (Steigerung des Energieumsatzes, Förderung der Glykogenspaltung, Fettmobilisierung), Kalzitonin (Kalziumhaushalt)")
263
Periphere Hormondrüsen
ENDOKRINOLOGIE Periphere Hormondrüsen NEBENSCHILDDRÜSEN PTH Parathormon (Parathyrin) für die Regulation des Kalziumhaushalts NEBENNIERENRINDE diese werden unter dem Überbegriff Kortikosteroide zusammengefasst In der äußeren Schicht der NNR (zona glomerulosa) erfolgt die Produktion der Mineralokortikoide (Aldosteron: wichtige Funktion bei der Regulation des Elektrolyt- und Flüssigkeitshaushalts; fördert die Rückresorption von Na in den Nieren)
 für die Regulation des Kalziumhaushalts. NEBENNIERENRINDE diese werden unter dem Überbegriff Kortikosteroide zusammengefasst. In der äußeren Schicht der NNR (zona glomerulosa) erfolgt die Produktion der Mineralokortikoide (Aldosteron: wichtige Funktion bei der Regulation des Elektrolyt- und Flüssigkeitshaushalts; fördert die Rückresorption von Na in den Nieren)")
264
Periphere Hormondrüsen
ENDOKRINOLOGIE Periphere Hormondrüsen NEBENNIERENRINDE In der mittleren Schicht (zona fasciculata) werden die Glukokortikoide (Kortisol) erzeugt. Kortisol beeinflusst den Kohlehydrat- und Eiweißstoffwechsel (Verstärkung der Glukoneogenese in der Leber) Verstärkung der Herzkraft sowie Gefäßverengung in der Peripherie (Verstärkung der Katecholamine) Entzündungshemmung (Einschränkung der unspezifischen und der spezifischen Immunabwehr)
 werden die Glukokortikoide (Kortisol) erzeugt. Kortisol beeinflusst den. Kohlehydrat- und Eiweißstoffwechsel (Verstärkung der Glukoneogenese in der Leber) Verstärkung der Herzkraft sowie Gefäßverengung in der Peripherie (Verstärkung der Katecholamine) Entzündungshemmung (Einschränkung der unspezifischen und der spezifischen Immunabwehr)")
265
Periphere Hormondrüsen
ENDOKRINOLOGIE Periphere Hormondrüsen NEBENNIERENRINDE In der inneren Schicht (zona reticularis) werden die Androgene (Testosteron) erzeugt. Allerdings ist die Bedeutung der Androgenbildung in der NNR nur sehr gering (~ 5% der testikulären Menge) DHEA Dehydroepiandrosteron NEBENNIERENMARK Die Zellen des NNM werden von sympathischen Nervenfasern des vegetativen NS versorgt; Katecholamine (Dopamin, Noradrenalin, Adrenalin) [Ausgangssubstanz für die Bildung ist die AS Tyrosin]
 werden die Androgene (Testosteron) erzeugt. Allerdings ist die Bedeutung der Androgenbildung in der NNR nur sehr gering (~ 5% der testikulären Menge) DHEA Dehydroepiandrosteron. NEBENNIERENMARK Die Zellen des NNM werden von sympathischen Nervenfasern des vegetativen NS versorgt; Katecholamine (Dopamin, Noradrenalin, Adrenalin) [Ausgangssubstanz für die Bildung ist die AS Tyrosin]")
266
Periphere Hormondrüsen
ENDOKRINOLOGIE Periphere Hormondrüsen NEBENNIERENMARK Das infolge eines Reizes aus dem NNM ausgeschüttete Hormongemisch enthält ~ 80% A, ~ 20% NA, ~ 2% DA A + NA wirken als Hormone und als Neurotransmitter über die Adrenorezeptoren (adrenerge Rezeptoren) an der Außenseite der Zellmembranen bzw. an den Endigungen sympathischer Nervenzellen NA + A werden auch als STRESSHORMONE (psychisch + physisch) bezeichnet
 an der Außenseite der Zellmembranen bzw. an den Endigungen sympathischer Nervenzellen. NA + A werden auch als STRESSHORMONE (psychisch + physisch) bezeichnet.")
267
Periphere Hormondrüsen
ENDOKRINOLOGIE Periphere Hormondrüsen Wirkungen der Katecholamine Weitstellung der Bronchien Weitstellung von Gefäßen in der Arbeitsmuskulatur Konstriktion von Gefäßen in der nicht-arbeitenden Muskulatur Steigerung der Hf und der Kontraktilität Hemmung der Magen-Darm Tätigkeit Förderung der Glykogenolyse (Leber und Muskel) Förderung der Lipolyse
 Förderung der Lipolyse.")
268
Periphere Hormondrüsen
ENDOKRINOLOGIE Periphere Hormondrüsen Inselorgan der BAUCHSPEICHELDRÜSE Insulin und Glukagon (beide sind am Kohlenhydratstoffwechsel beteiligt) Speicherung der Nahrung in Form von Glykogen und Fett (Insulin!) Mobilisation der Energiedepots bei Arbeit (Glukose) Konstanthaltung des Blutglukosespiegels (Ins, Gluk) Förderung des Wachstums (Ins)
 Speicherung der Nahrung in Form von Glykogen. und Fett (Insulin!) Mobilisation der Energiedepots bei Arbeit (Glukose) Konstanthaltung des Blutglukosespiegels. (Ins, Gluk) Förderung des Wachstums (Ins)")
269
Periphere Hormondrüsen
ENDOKRINOLOGIE Periphere Hormondrüsen KEIMDRÜSEN Die Produktion der Sexualhormone (von der chemischen Struktur her handelt es sich um Steroide) wird über das hypothalamische GnRH, und über die adenohypophysären Hormone FSH und LH geregelt Androgene (männliche Sexualhormone) werden in den Testes (Leydig‘schen Zwischenzellen) gebildet; geringe Androgenmengen werden auch bei der Frau in den Ovarien und in der NNR gebildet
 wird über das hypothalamische GnRH, und über die adenohypophysären Hormone FSH und LH geregelt. Androgene (männliche Sexualhormone) werden in den Testes (Leydig‘schen Zwischenzellen) gebildet; geringe Androgenmengen werden auch bei der Frau in den Ovarien und in der NNR gebildet.")
270
Periphere Hormondrüsen
ENDOKRINOLOGIE Periphere Hormondrüsen KEIMDRÜSEN Östrogene (Östron, Östradiol) und Gestagene (Progesteron) (weibliche Sexualhormone) werden hauptsächlich in den Ovarien gebildet. Beim Mann werden geringe Mengen an Östrogenen und Gestagenen in den Hoden gebildet Die Hauptwirkung dieser Hormone wird v.a. in der Pubertät augenfällig (androgene und anabole Wirkung des Testosteron beim Mann; „Vermännlichung des Körpers“ - bzw. die gynoide Wirkung bei der Frau: Eireifung, Menstruationszyklus, „Verweiblichung“ des Körpers)
 und Gestagene (Progesteron) (weibliche Sexualhormone) werden hauptsächlich in den Ovarien gebildet. Beim Mann werden geringe Mengen an Östrogenen und Gestagenen in den Hoden gebildet. Die Hauptwirkung dieser Hormone wird v.a. in der Pubertät augenfällig (androgene und anabole Wirkung des Testosteron beim Mann; „Vermännlichung des Körpers - bzw. die gynoide Wirkung bei der Frau: Eireifung, Menstruationszyklus, „Verweiblichung des Körpers)")
271
Auswirkungen von Training auf das endokrine System
ENDOKRINOLOGIE Auswirkungen von Training auf das endokrine System intensives Ausdauertraining über längere Zeiträume führt dazu, dass bei identischer Belastung (in Absolut-Werten) der Hormonanstieg geringer ausfällt (z.B. Katecholamine, HGH) bei maximaler Belastung weisen Hochausdauertrainierte eher höhere Hormonspiegel auf als Untrainierte
 der Hormonanstieg geringer ausfällt (z.B. Katecholamine, HGH) bei maximaler Belastung weisen Hochausdauertrainierte eher höhere Hormonspiegel auf als Untrainierte.")
272
Auswirkungen von Training auf das endokrine System
ENDOKRINOLOGIE Auswirkungen von Training auf das endokrine System Für Insulin zeigt sich bei Trainierten eine erhöhte Sensitivität an den Zielzellen (v.a. Muskelgewebe) Die Testosteronwerte sind bei krafttrainierten Sportlern höher als ‚Normalwerte‘ bei Untrainierten-und dort wiederum etwas höher als bei Ausdauertrainierten Der Einfluss von Leistungssport (v.a. extreme Langzeitausdauer) wirkt sich bei Frauen oftmals ‚negativ‘ aus-und beeinträchtigt die gonadale Achse (verstärkt bei kalorienreduzierter Diät und geringem Körperfettanteil)
 Die Testosteronwerte sind bei krafttrainierten Sportlern höher als ‚Normalwerte‘ bei Untrainierten-und dort wiederum etwas höher als bei Ausdauertrainierten. Der Einfluss von Leistungssport (v.a. extreme Langzeitausdauer) wirkt sich bei Frauen oftmals ‚negativ‘ aus-und beeinträchtigt die gonadale Achse (verstärkt bei kalorienreduzierter Diät und geringem Körperfettanteil)")
273
Grundlagen der Körperzusammensetzung
(Body Composition) Theorie und Praxis
 Theorie und Praxis.")
274
Inhalt Überblick über Body Composition und Methoden
Theoretischer Hintergrund Anwendungsmöglichkeiten

275
Warum wird eine Körperfettmessung durchgeführt ?
Übergewicht Adipositas Beziehung zwischen dem BMI (kg/m2) und dem Morbiditäts- und Mortalitätsrisiko Metropolitan Life Insurance ~ 4.9 Mill. Policen von „Gewicht mit der geringsten Sterblichkeit“ [Build and Blood Pressure Study, Society of Actuaries 1959] 1983 revidiert => Idealgewicht liegt höher [Build Study, Society of Actuaries 1979]
 und dem Morbiditäts- und Mortalitätsrisiko. Metropolitan Life Insurance. ~ 4.9 Mill. Policen von „Gewicht mit der geringsten Sterblichkeit [Build and Blood Pressure Study, Society of Actuaries 1959] 1983 revidiert => Idealgewicht liegt höher. [Build Study, Society of Actuaries 1979]")
276
ADIPOSITAS (adiposity, obesity): erhöhter Anteil des Körperfettes an der Körpermasse
ÜBERGEWICHT (overweight): erhöhte Körpermasse im statistischen Mittel (keine Aussage darüber, wie und woraus sich ein erhöhtes KG ‚zusammensetzt‘) Somit kann jemand adipös sein ohne übergewichtig zu sein, und jemand kann übergewichtig sein ohne notwendigerweise adipös zu sein
: erhöhte Körpermasse im statistischen Mittel (keine Aussage darüber, wie und woraus sich ein erhöhtes KG ‚zusammensetzt‘) Somit kann jemand adipös sein ohne übergewichtig zu sein, und jemand kann übergewichtig sein ohne notwendigerweise adipös zu sein.")
277
BMI 20-25 = Grad 0 (Normalgewicht)
„Normwertangaben“ aus der Literatur: Als Grenze zur Fettleibigkeit Männer > 25% GKF, Frauen > 30% GKF [nach Bray 1987] Body Mass Index (BMI) > 30: „....deutlicher Anstieg der Mortalität“ [Garrow 1988] BMI = Grad 0 (Normalgewicht) BMI = Grad I (Übergewicht) BMI = Grad II (Adipositas) BMI > 40 = Grad III (morbide A.) [Garrow 1988, NIH Consensus Development Conference 1985]
 > 30: „....deutlicher Anstieg der Mortalität [Garrow 1988] BMI = Grad 0 (Normalgewicht) BMI = Grad I (Übergewicht) BMI = Grad II (Adipositas) BMI > 40 = Grad III (morbide A.) [Garrow 1988, NIH Consensus Development Conference 1985]")
278
Direkter Zusammenhang zwischen dem Grad der Adipositas bzw
Direkter Zusammenhang zwischen dem Grad der Adipositas bzw. der Fettverteilung und dem Morbiditätsrisiko bzw. Mortalitätsrisiko viszerale Fettgewebe ist das gefährliche (Bauchfett) [inverse J-shape] BMI > 35 BMI 30-35 Gesundheitsrisiko BMI 26-29 BMI 22-25 Body Mass Index
 [inverse J-shape] BMI > 35. BMI Gesundheitsrisiko. BMI BMI Body Mass Index.")
280
Gibt es sowas wie Normwerte (sehr gut, gut, durchschnittlich, schlecht) für das GKF ?
Statistisch betrachtet JA; biologisch-medizinisch betrachtet JEIN Ein deutliches ZUVIEL an KF ist genauso problematisch wie ein deutliches ZUWENIG GKF ist abhängig von: (i) Alter, (ii) Geschlecht, (iii) Trainingszustand bzw. Grad der körperl. Aktivität, (iv) Ernährungsgewohnheiten (soziales Umfeld), (v) Genetik
 Alter, (ii) Geschlecht, (iii) Trainingszustand bzw. Grad der körperl. Aktivität, (iv) Ernährungsgewohnheiten (soziales Umfeld), (v) Genetik.")
281
bei Männern GKF <10% = trainierte M. bzw. Jugendliche GKF 10-15% = durchschnittlicher, schlanker Mann GKF bis ~ 20% = typischer Mann GKF > 20 = Hände weg vom Bier bei Frauen GKF <15% = trainierte Frau GKF 15-20% = durchschnittliche, schlanke Frau GKF 20-25% = typische Frau GKF > 25 (30)% = weniger oft Konditorei
% = weniger oft Konditorei.")
282
Normwerte für GKF international
Tab 1.: Idealer Körperfettanteil in Abhängigkeit von Alter und Geschlecht Normwerte für GKF international Frauen Körperfett in % Alter Männer Körperfett in % 18,9 22,1 25,0 29,6 20-24 10,8 14,9 19,0 23,3 22,0 25,4 29,8 25-29 12,8 16,5 20,3 24,3 19,7 22,7 26,4 30,5 30-34 14,5 18,0 21,5 25,2 21,0 24,0 27,7 31,5 35-39 16,1 19,3 22,6 26,1 25,6 29,3 32,8 40-44 17,5 20,5 23,6 26,9 27,3 30,9 34,1 45-49 18,6 24,5 27,6 25,8 28,9 32,3 35,5 50-54 19,5 22,3 28,3 27,0 30,2 33,5 36,7 55-59 20,0 22,9 25,9 34,2 37,7 60 + 23,4 29,5

283
Warum BCA ? Einige Anwendungsmöglichkeiten….
Abschätzung möglicher gesundheitlicher Risikofaktoren (zuviel/zu wenig Körper-fett) Erfassung der Änderung der BC im Zuge von Training und/oder spezieller Ernährungsformen Erfassung der Änderung der BC im Zuge der natürlichen biologischen Entwicklung (Wachstum, altersbedingte Änderungen der BC…)
 Erfassung der Änderung der BC im Zuge von Training und/oder spezieller Ernährungsformen. Erfassung der Änderung der BC im Zuge der natürlichen biologischen Entwicklung (Wachstum, altersbedingte Änderungen der BC…)")
284
zur Fettverteilung.... Fettverteilung mit verschiedenen Erkrankungen assoziiert (J Vague, 1954/1956) ...der androide Typ ...der gynoide Typ

285
Körperfettverteilung
2 wesentliche Körperfettdepots ….das subkutane Körperfettgewebe (‚Speicherfettgewebe‘) das sich fast am gesamten Körper anlegen kann und als Energiespeicher, Schutzfaktor, Modulator verschiedener Hormone etc. dient ….das viszerale (intrabadominale) Fettgewebe (‚Bierbauch‘) das sich im Laufe des Lebens anlagert bzw die Folge von hormonellen Schwankungen sein kann
 das sich fast am gesamten Körper anlegen kann und als Energiespeicher, Schutzfaktor, Modulator verschiedener Hormone etc. dient. ….das viszerale (intrabadominale) Fettgewebe (‚Bierbauch‘) das sich im Laufe des Lebens anlagert bzw die Folge von hormonellen Schwankungen sein kann.")
286
Das subkutane Fettgewebe stellt eher ein kosmetisches Problem dar
Körperfettverteilung Metabolisch und biochemisch betrachtet sind beide Fettdepots sehr unterschiedlich Das subkutane Fettgewebe stellt eher ein kosmetisches Problem dar Das viszerale Fettgewebe ist das gesundheitlich ‚bedenklichere‘ (erhöht den Anteil an FFS im Splanchnikusgebiet und kann in weiterer Folge zu den bekannten Erscheinungsformen, wie Typ II-Diabetes, Herzinfarkt und Schlaganfall führen)
")
287
Wie kann das innenliegende KF relativ einfach
Körperfettverteilung Wie kann das innenliegende KF relativ einfach ‚gemessen‘ werden ? Anthropometrischer Ansatz: mit dem Massband wird der Bauchumfang (in cm) gemessen bzw. zusätzlich der Hüftumfang (in cm). Über den Vergleich mit Referenzwerten (nach Alter und Geschlecht) erfolgt dann eine „Risikoabschätzung“
 gemessen bzw. zusätzlich der Hüftumfang (in cm). Über den Vergleich mit Referenzwerten (nach Alter und Geschlecht) erfolgt dann eine „Risikoabschätzung")
288
♀ WHR ~ 0.70-0.80 [ 0.85] ♂ WHR ~ 0.80-0.85 (0.90) [ 0.95]
Körperfettverteilung Entweder wird nur der BU für die Interpretation herangezogen (BU zeigt normalerweise sehr gute positive Korrelation mit dem viszeralen Fettanteil) oder das Verhältnis von BU zu HU („Waist-to-Hip-Ratio, WHR“) wird für die Abschätzung eines gesundheitlichen Risikos herangezogen ♀ WHR ~ [ 0.85] ♂ WHR ~ (0.90) [ 0.95]
![♀ WHR ~ [ 0.85] ♂ WHR ~ (0.90) [ 0.95]](http://slideplayer.org/slide/874998/2/images/288/%E2%99%80+WHR+%7E+%5B%EF%82%B3+0.85%5D+%E2%99%82+WHR+%7E+%280.90%29+%5B%EF%82%B3+0.95%5D.jpg "Körperfettverteilung. Entweder wird nur der BU für die Interpretation herangezogen (BU zeigt normalerweise sehr gute positive Korrelation mit dem viszeralen Fettanteil) oder das Verhältnis von BU zu HU („Waist-to-Hip-Ratio, WHR ) wird für die Abschätzung eines gesundheitlichen Risikos herangezogen. ♀ WHR ~ [ 0.85] ♂ WHR ~ (0.90) [ 0.95]")
289
„At the population level, measurement of body
Körperfett und Risiko??? „At the population level, measurement of body FM has no advantage over BMI and WC in the prediction of obesity-related metabolic risk. Although measures of central adiposity (WC, WC/ht) tended to show closer associations with risk factors than measures of general adiposity, the differences were small and depended on the type of risk factor and sex, suggesting an equivalent value of methods.“ Bosy-Westphal et al.: Value of body fat mass vs anthropometric obesity indices in the assessment of metabolic risk factors. IJO Nov 2005
 tended to show closer associations with risk factors. than measures of general adiposity, the differences. were small and depended on the type of risk factor and sex, suggesting an equivalent value of methods. Bosy-Westphal et al.: Value of body fat mass vs anthropometric obesity indices in the assessment of metabolic risk factors. IJO Nov")
290
Aufbau des Körpers und Modelle der Körperzusammensetzung
Body Composition Körperfettverteilung Wo befindet sich das Körperfett und auch wieviel ist wo vorhanden (Topologie) Primär quantitativ orientierter Ansatz; woraus setzen sich einzelne Körper-kompartments zusammen
 Primär quantitativ orientierter Ansatz; woraus setzen sich einzelne Körper-kompartments zusammen.")
291
KF KF KF Wasser EZF KF- freien Anteil IZF Protein IZS Mineral EZS
Body Composition Für die praktische Arbeit und Messung haben sich die sog. Komponenten (od. Kompartment-) modelle bewährt Flüssig-Metabol.Modell KF KF KF Wasser EZF KF- freien Anteil IZF Protein IZS Mineral 2K-Modell EZS 4K-(chem)Modell
 modelle bewährt. Flüssig-Metabol.Modell. KF. KF. KF. Wasser. EZF. KF- freien. Anteil. IZF. Protein. IZS. Mineral. 2K-Modell. EZS. 4K-(chem)Modell.")
292
Mineral Fettgewebe Protein Weiches Gewebe nicht Musk. LFM FFM Wasser
Body Composition Mineral Fettgewebe Protein Weiches Gewebe nicht Musk. LFM FFM Wasser Muskel gewebe Ess.Lip Knochen Lipid Fett Nicht-Ess.Lip 4K-(anatom)Modell
Modell.")
293
Männer

294
Frauen

296
Einige der gängigsten Messsmethoden
Body Composition Einige der gängigsten Messsmethoden 1-Kompartment Methoden: Waage (de facto nur das Kompartment Körpermasse) 2-Kompartment Methoden: alle Methoden, die entweder Körperfett direkt/indirekt messen (können) oder die fettfreie Körpermasse direkt/indirekt bestimmen (können) {Kaliper, Lipometer, Bildgebende Verfahren, Futrex(?), Unterwasserwiegen (?)}
 2-Kompartment Methoden: alle Methoden, die entweder Körperfett direkt/indirekt messen (können) oder die fettfreie Körpermasse direkt/indirekt bestimmen (können) {Kaliper, Lipometer, Bildgebende Verfahren, Futrex( ), Unterwasserwiegen ( )}")
297
Kaliper (Anwendung, Vorteile, Nachteile)
")
298
Body Composition Messsmethoden 3-Kompartment Methoden: alle Methoden, die zB. Flüssigkeitsräume direkt/indirekt messen (können), od. den Wechselstromwiderstand des Körpers messen (können) {K40-Bestimmung, Deuterium-Dilution, Bioimpedanz (bipolar, tetrapolar), Dual Energy X-Ray Absorptiometry(DEXA)}
, od. den Wechselstromwiderstand des Körpers messen (können) {K40-Bestimmung, Deuterium-Dilution, Bioimpedanz (bipolar, tetrapolar), Dual Energy X-Ray Absorptiometry(DEXA)}")
299
BIA Bioelectrical Impedance Analysis
BIA misst den Widerstand, den der Körper einem angelegten Wechselstrom entgegensetzt

300
BIA

301
Zum Phasenwinkel

302
Multifrequenz vs. Singlefrequenz

303
BIA BIA Vektor

304
BIA Vektor bei einem Triathleten

305
Eingesetzt für Messungen der Knochendichte (Osteoporose-HRT)
DEXA Eingesetzt für Messungen der Knochendichte (Osteoporose-HRT)
")
306
DEXA DEXA auch für Körperfettmessungen eingesetzt (‚Golden Standard‘) [Problem: Softwareversionen, Annahmen, starke Annäherung der %KF an den BMI)
 [Problem: Softwareversionen, Annahmen, starke Annäherung der %KF an den BMI)")
307
Tobec

308
Tobec

Ähnliche Präsentationen
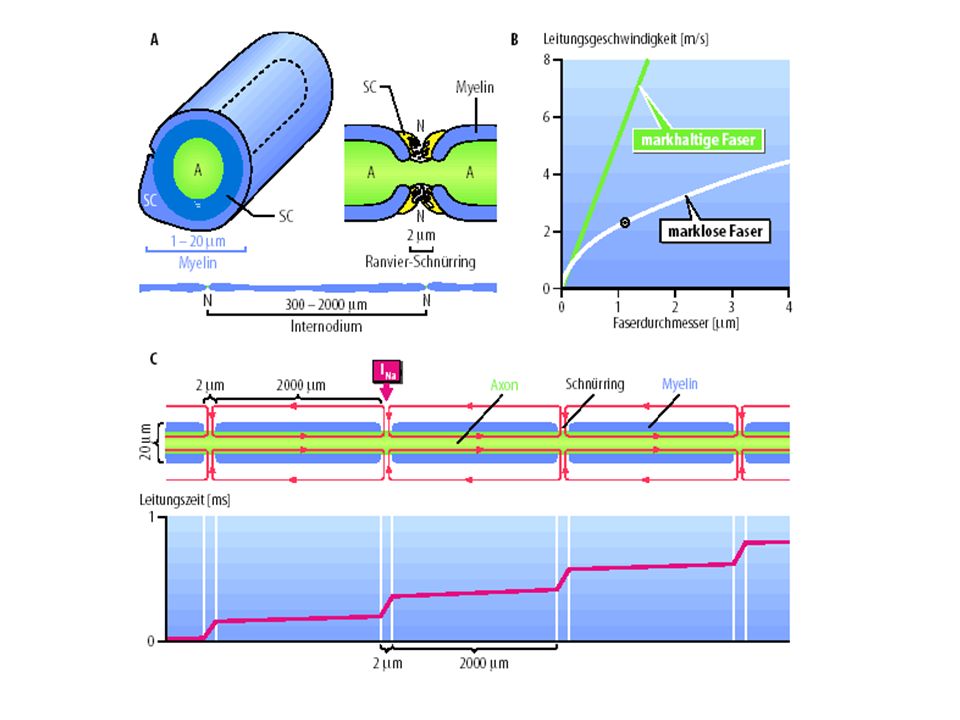

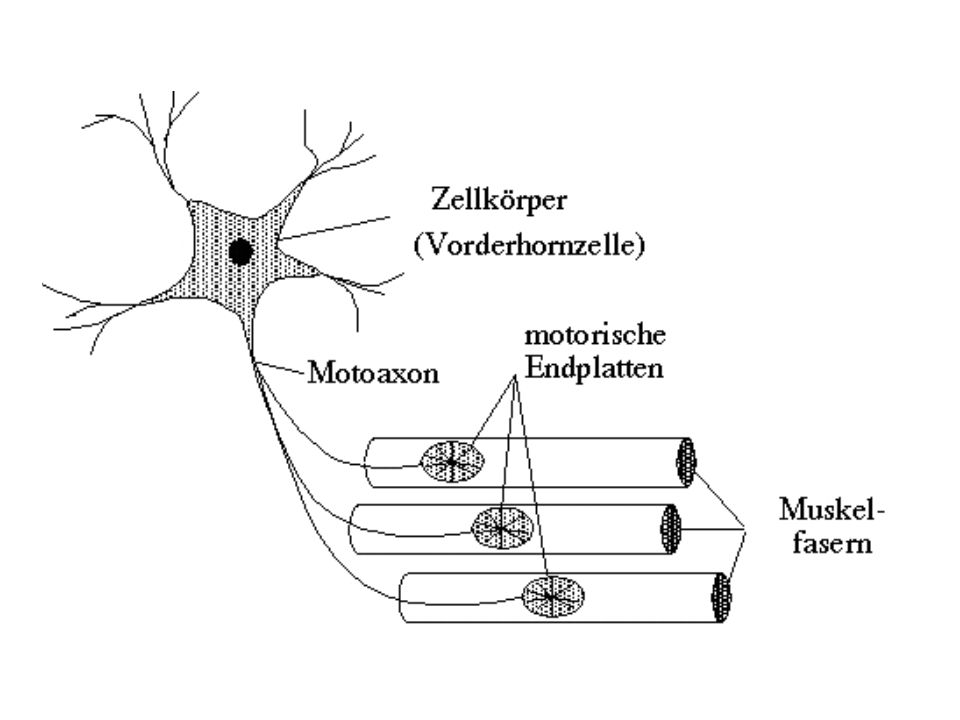
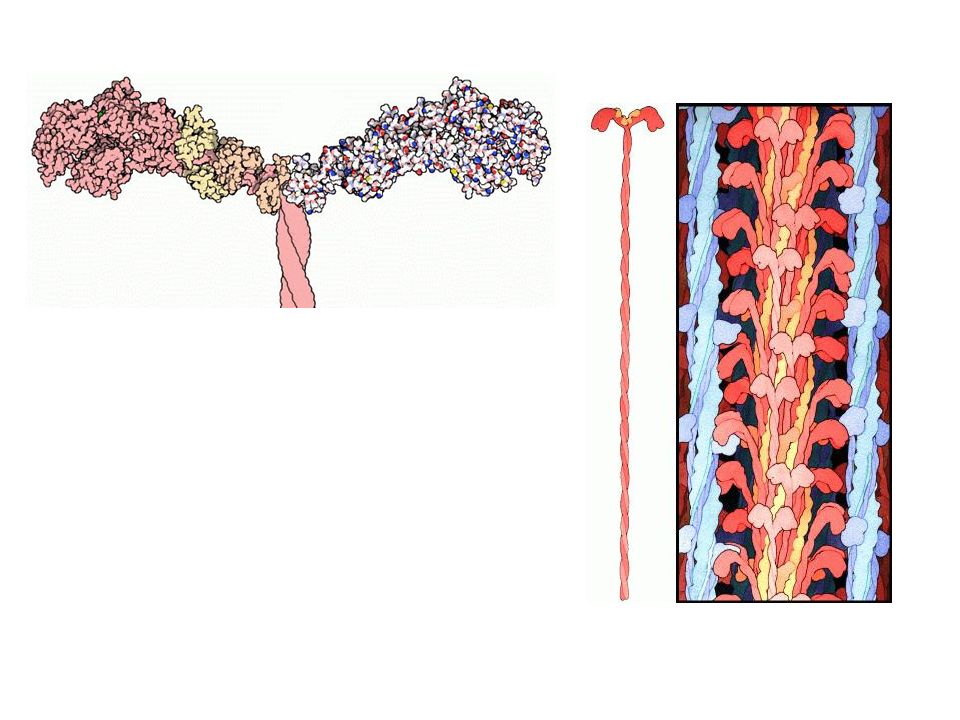
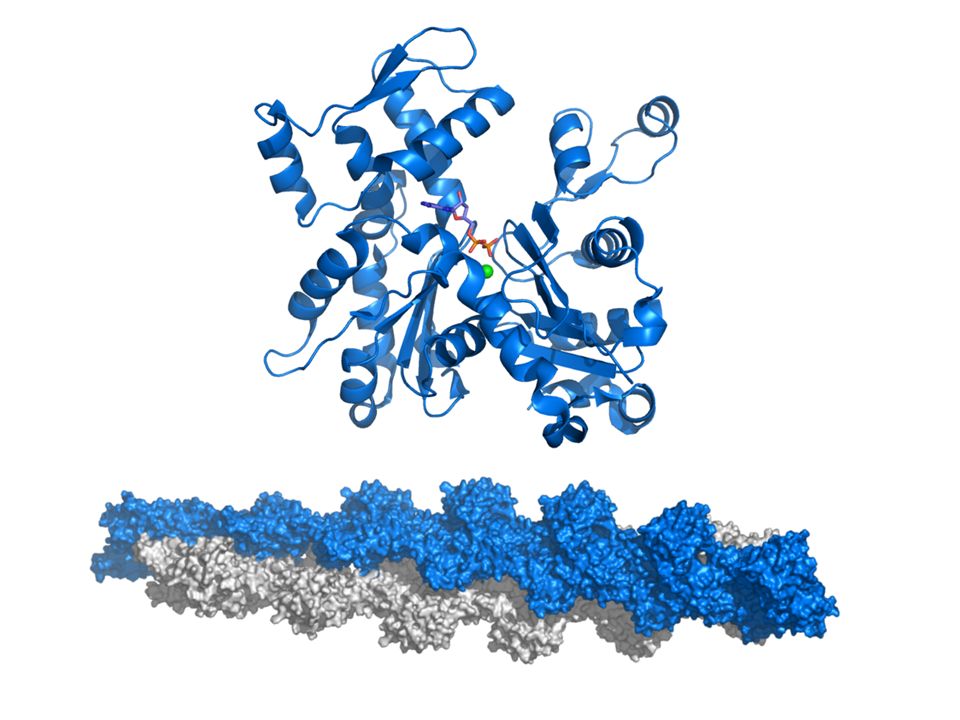
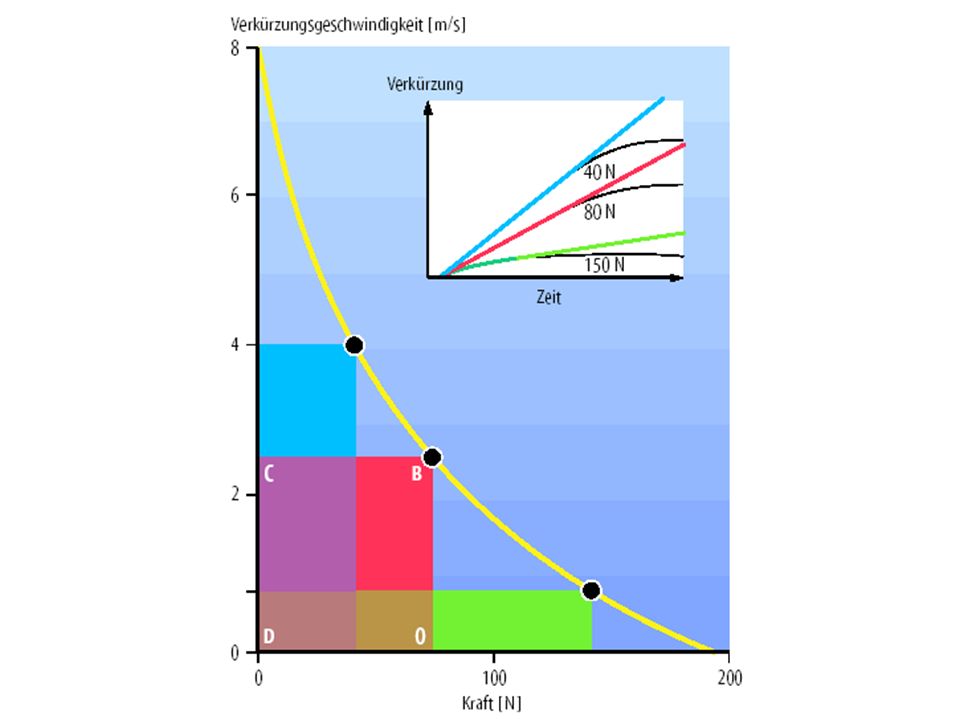
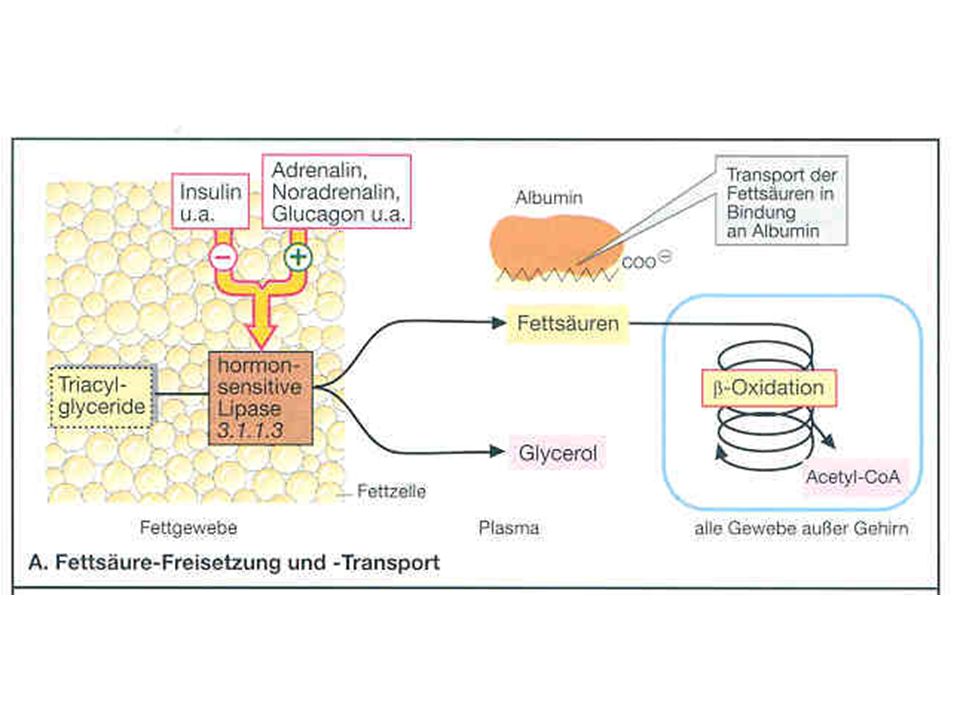
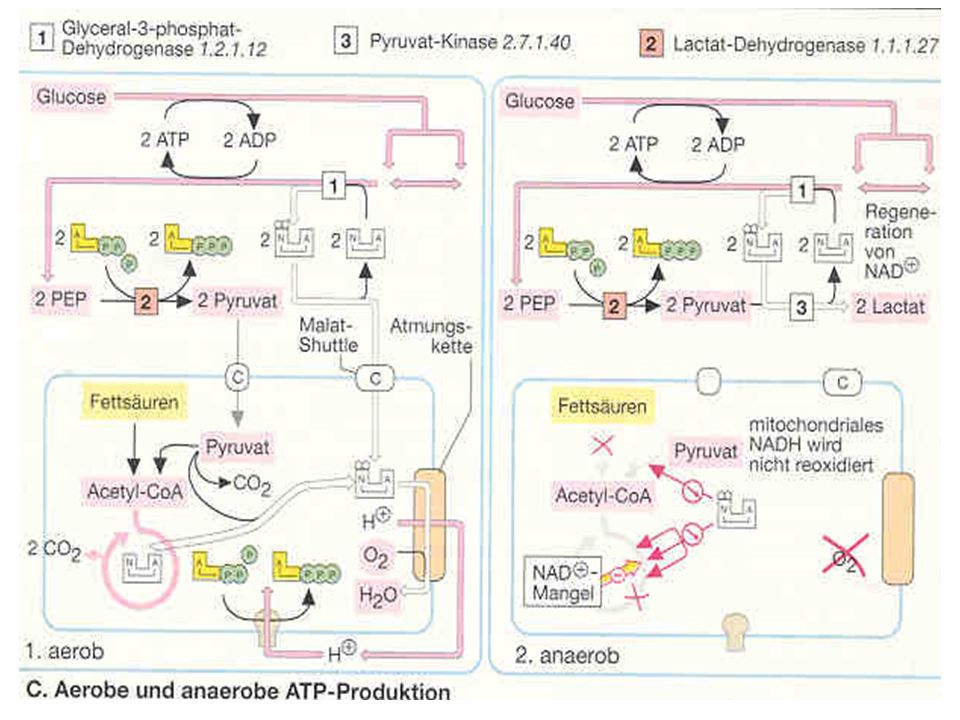
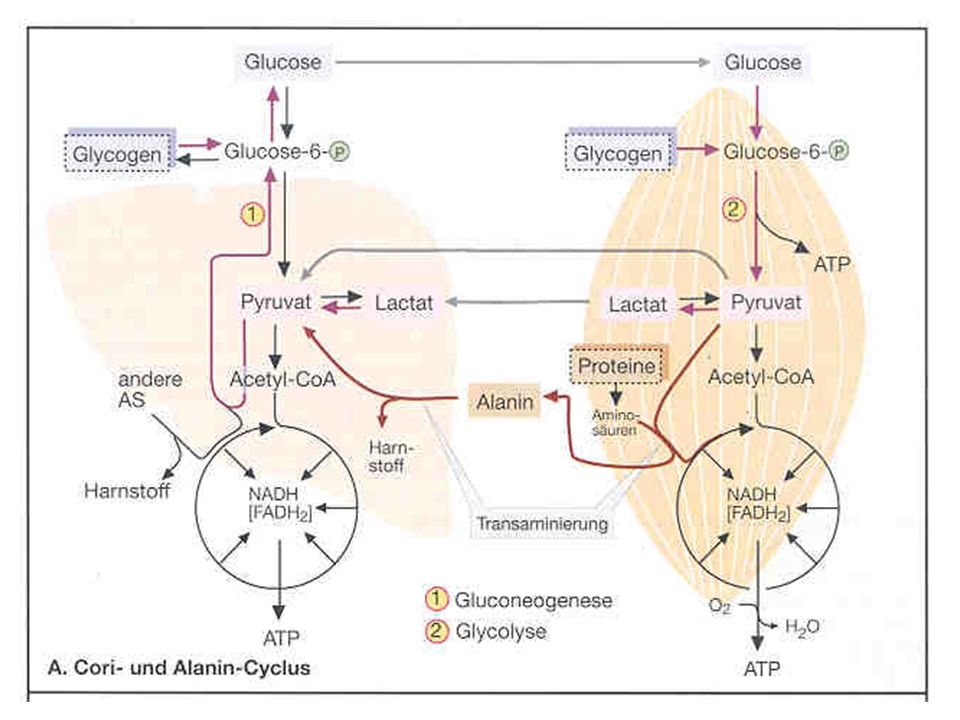

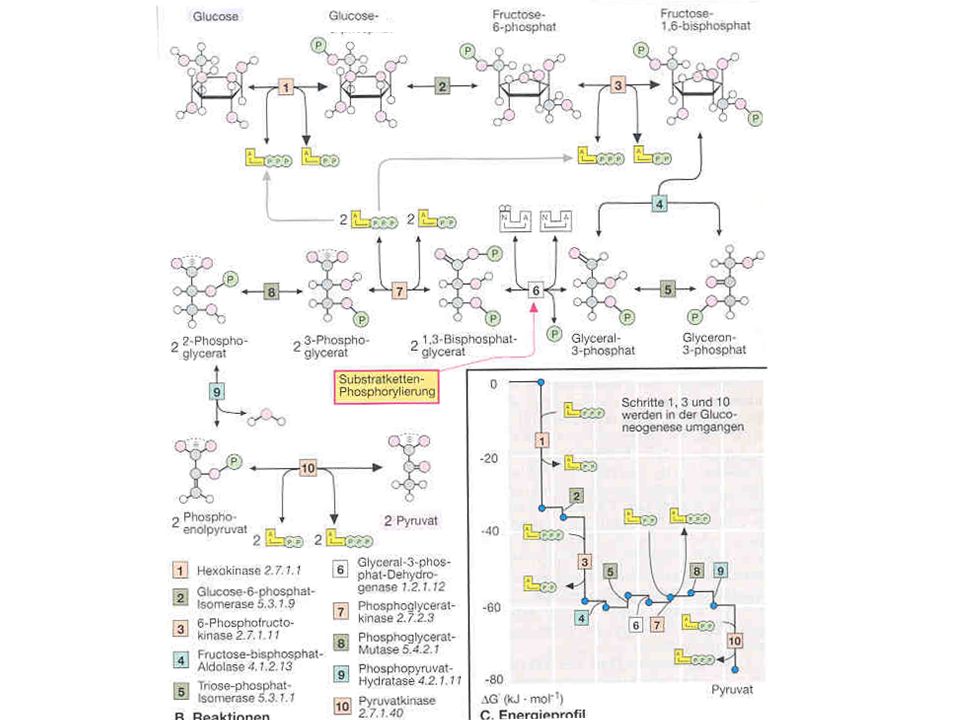
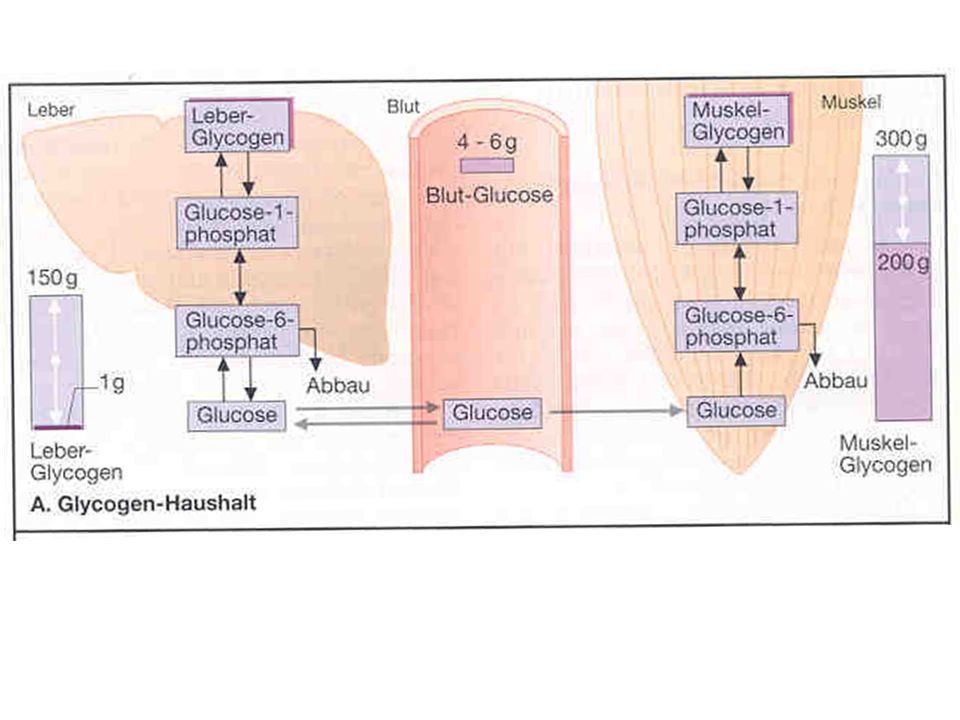



>")





>")









 Glucose.>")


