Präsentation herunterladen
Die Präsentation wird geladen. Bitte warten

1
Tumorbedingte Gefäßneubildung
Differentialgleichungen in der Biomedizin SoSe 09 Philipp Schmauck

2
Avaskuläre Tumore: Avaskuläre Tumore:
Nekrotischer Kern aufgrund von Nährstoffmangel Zwischenschicht aus ruhigen Zellen Außenschicht aus sich vermehrenden Zellen Gleichgewicht zwischen Mitose, Apoptose und der Auflösung von Tumorzellen in Abfallstoffe Tumor ist in seiner Größe beschränkt

3
Avaskuläre Tumore: Für weiteres Wachstum und Metastasierung benötigt der Tumor die Nährstoffversorgung durch einen Blutkreislauf Angiogenese: Wachstum von Kapillaren durch Sprossung aus einem bestehenden Kapillarsystem Endothelzellen an der Innenseite des Blutgefäßes spielen hierbei eine wichtige Rolle

4
Basement Membrane (BM/BL)
Fibroblast Capillary Endothelial cells (EC) Extracellular Matrix(ECM)
 Extracellular Matrix(ECM)")
5
EC werden stimuliert proteolytische Enzym auszuschütten
1. Tumor sondert angiogenetische Wachstumsfaktoren ab , hier Vascular Endothelial Growth Factor (VEGF) EC werden stimuliert proteolytische Enzym auszuschütten Enzym steuert Abbau BM EC durchdringen BM und migrieren in Richtung Quelle des VEGF Neue Kapillaren entstehen durch Proliferation (Vermehrung) und Migration (Wanderung) Es entsteht ein Kapillar-Netzwerk Dies geschieht bis das Kapillar-Netzwerk den Tumor erreicht, in ihn eindringt und ihn mit Nährstoffen versorgt Kapillare Tumor 2.
 EC werden stimuliert proteolytische Enzym auszuschütten. Enzym steuert Abbau BM. EC durchdringen BM und migrieren in Richtung Quelle des VEGF. Neue Kapillaren entstehen durch Proliferation (Vermehrung) und Migration (Wanderung) Es entsteht ein Kapillar-Netzwerk. Dies geschieht bis das Kapillar-Netzwerk den Tumor erreicht, in ihn eindringt und ihn mit Nährstoffen versorgt. Kapillare. Tumor. 2.")
6
Geometrie des Problems
G(x,y,t) g(x,t)
 g(x,t)")
7
Biochemische Kinetik V + R ⇌ RV (k1, k-1) – Bindung VEGF (V) an EC Rezeptoren (R) RV → C + R (k2) – Produktion Proteolytische Enzym (C) und neuer Rezeptor (R) C + F → CF (k3) – Bindung Enzym an BM Rezeptoren (F) CF → F´ + C (k4) – Abbau der BM und Bildung Katalysator (F`) Fibronektin ist hier ein genereller Term für extrazelluläre Proteine
 – Produktion Proteolytische Enzym (C) und neuer Rezeptor (R) C + F → CF (k3) – Bindung Enzym an BM Rezeptoren (F) CF → F´ + C (k4) – Abbau der BM und Bildung Katalysator (F`) Fibronektin ist hier ein genereller Term für extrazelluläre Proteine.")
8
Anwendung Massenwirkungsgesetz
x – Position an der Kapillarwand t – Zeit v – Konzentration des angiogenetische Faktor V r – Dichte der Rezeptoren R auf den EC l - Konzentration des Rezeptor-Komplexes RV n – Konzentration von EC f – Konzentration von Fibronektin

9
Anwendung Massenwirkungsgesetz
Anwendung der MM-Kinetik auf 1. und 2. ergibt: Für 1. und 2.

10
Anwendung MM-Kinetik Anwendung der MM-Kinetik auf 3. und 4. ergibt:
Für 3. und 4.

11
Zusätzliche Bedingungen
Proteolytische Enzym zerfällt proportional zu seiner Konzentration Zerfallskonstante EC produzieren Fibronektin Logistische Funktion 2. Term ist ein logistische Differentialgleichung

12
Anfangsbedingungen l(x,0) = 0 - Am Anfang existiert kein Rezeptor-Komplex c(x,0) ≈ 0 - Am Anfang sind wenig proteolytische Enzyme vorhanden f(x,0)=fM(x) – Fibronektin Anfangswert ist gleich dem Wert in normalen Zellen v(x,0) – kann von uns beliebig vorgegeben werden Bestimmung von n(x,0) r(x,0) problematisch
=fM(x) – Fibronektin Anfangswert ist gleich dem Wert in normalen Zellen. v(x,0) – kann von uns beliebig vorgegeben werden. Bestimmung von n(x,0) r(x,0) problematisch.")
13
Anfangsbedingungen

14
Anfangsbedingungen Annähernd konstant und der Wert ist relativ einfach zu ermitteln: Durchmesser Kapillare: 6-8 µM Durchmesser rote Blutkörperchen: 4-5 µM Dann können wir abschätzen: Dicke der EC 1 µM und Breite 10 µM Vernachlässigung der Dicke der BM Existieren EC pro mm D.h. Länge der EC: µM D.h. die volumenbezogene Dichte der EC: Zellen pro Liter Anzahl der Rezeptoren pro Zelle ist von der Ordnung: 105

15
Anfangsbedingungen Und wir können schreiben:

16
Bewegung der EC Kapillarwand ist eindimensionales Gitter
EC sind gleichverteilt, berühren sich nicht und sind angeordnet an Referenzpunkt nh W - Kontrollsubstanz τ´n± (W) - Wahrscheinlichkeit eines Schrittes einer EC von n zu n+1, n-1 nn(t) - Wahrscheinlichkeitsdichte der Verteilung der EC an Position n zur Zeit t Berücksichtigung einer Wartezeit
 - Wahrscheinlichkeit eines Schrittes einer EC von n zu n+1, n-1. nn(t) - Wahrscheinlichkeitsdichte der Verteilung der EC an Position n zur Zeit t. Berücksichtigung einer Wartezeit.")
17
Bewegung der EC Änderung von nn(t):
Teilchen die von (n±1)h nach nh hinzu wandern Teilchen die von nh nach (n±1)h abwandern Erwartete Wartezeit eines Teilchens in n bis es n wieder verlässt: Wahrscheinlichkeitsdichte ist gleich der Dichte der EC; durchschnittliche Wartezeit ist gleich Erwartungswert geometrische Verteilung
h nach nh hinzu wandern. Teilchen die von nh nach (n±1)h abwandern. Erwartete Wartezeit eines Teilchens in n bis es n wieder verlässt: Wahrscheinlichkeitsdichte ist gleich der Dichte der EC; durchschnittliche Wartezeit ist gleich Erwartungswert geometrische Verteilung.")
18
Bewegung der EC Kontrollsubstanz beinhaltet die Effekte von VEGF auf die Zellen: W=(…,W-n-1/2, W-n, W-n+1/2,…) Wn=Wn(c,f) c – proteolytische Enzym: Abbau BM f – Fibronektin: Bestandteil BM
 c – proteolytische Enzym: Abbau BM. f – Fibronektin: Bestandteil BM.")
19
Bewegung der EC Annahme: Entscheidung „when to move“ ist unabhängig von der Entscheidung „ where to move“. D.h. Wartezeit in n ist konstant: Annahme: τ± hängt nur von benachbarten Kontrollsubstanzen ab:

20
Bewegung der EC

21
Taylorentwicklung

22
Taylorentwicklung

23
Bewegung der EC Setze τ(W(f,c))=τ1(c)τ2(f) – Auswirkung von Protease und Fibronektin auf EC: EC wandern in Gebiete mit hoher Protease Konzentration EC wandern in Gebiete mit geringer Fibronektin Konzentration Vermeidung von Singularität (ln(τ) und Ableitung):
 und Ableitung):")
24
Numerische Simulation

25
Numerische Simulation
2 1 (1-cos(2pix)) ist größer 0 und kleiner 2 und für x€[0,1] genau eine Spitze
) ist größer 0 und kleiner 2 und für x€[0,1] genau eine Spitze.")
26
Numerische Simulation
n(x,t) D=3,6*10-5 α1=0,001 α2=1,0 γ1=1,2 n β1=1,0 β2=0,001 γ2=1,2 v λ1=73,0 υ1=0,007 m=100 ν0=15 c f β=0,222 λ2=19,0 v1=1,28
 D=3,6*10-5. α1=0,001. α2=1,0. γ1=1,2. n. β1=1,0. β2=0,001. γ2=1,2. v. λ1=73,0. υ1=0,007. m=100. ν0=15. c. f. β=0,222. λ2=19,0. v1=1,28.")
27
Numerische Simulation
Unmittelbarer Fibronektin Abbau in 0,44 < x < 0,56 Abbau ca. Kapillar Durchmesser von ~6μM

28
Numerische Simulation
EC Bewegung Andeutung Kapillare Sprossung

29
Numerische Simulation
Höchste Konzentration in 0,44 < x < 0,56 Rapide Abnahme des Wachstumfaktors

30
Numerische Simulation
Proteolytische Enzyme konvergieren zu „steady-sate“

31
Angiostatin Angiogenese Hemmer: Natürliches Protein
Hemmt Bildung neuer Blutgefäße Direkter Hemmstoff für Protease Angiostatin stimuliert EC zur Produktion eines Hemmstoffes Klinische Untersuchung für die Krebstherapie

32
Biochemische Kinetik V + R ⇌ RV (k1, k-1) – Bindung VEGF (V) an EC Rezeptoren (R) RV → C + R (k2) – Produktion Proteolytische Enzym (C) und neuer Rezeptor (R) Direkter Inhibitor: A + CA ⇌ CI – Proteolytische Enzyme (CI) gehemmt vom Angiostatin (A) und Fibronektin abbauende Enzyme (CA) A ist Katalysator für Fibronektin Abbau
 – Produktion Proteolytische Enzym (C) und neuer Rezeptor (R) Direkter Inhibitor: A + CA ⇌ CI – Proteolytische Enzyme (CI) gehemmt vom Angiostatin (A) und Fibronektin abbauende Enzyme (CA) A ist Katalysator für Fibronektin Abbau.")
33
Biochemische Kinetik [CI]=ve[A][CA] – Indirekter Inhibitor:
A + RA ⇌ ARA (k3,k-3) – Rezeptor Protein (RA) auf EC bindet mit Angiostatin ARA → I+RA (k4) – Protease Inhibitor (I) produziert von EC in Reaktion auf Angiostatin [CI]=ve[A][CA] 3.1 ist im Gleichgewicht
![Biochemische Kinetik [CI]=ve[A][CA] – Indirekter Inhibitor:](http://slideplayer.org/slide/854862/2/images/33/Biochemische+Kinetik+%5BCI%5D%3Dve%5BA%5D%5BCA%5D+%E2%80%93+Indirekter+Inhibitor%3A.jpg "A + RA ⇌ ARA (k3,k-3) – Rezeptor Protein (RA) auf EC bindet mit Angiostatin. ARA → I+RA (k4) – Protease Inhibitor (I) produziert von EC in Reaktion auf Angiostatin. [CI]=ve[A][CA] 3.1 ist im Gleichgewicht.")
34
Biochemische Kinetik CA + F ⇌ CAF (k5, k-5) – Bindung Enzym an Fibronektin Rezeptoren (F) CAF → CA + F´ (k6) - Abbau Fibronektin und Bildung Katalysator (F`) [C]=[CA]+[CAF]+[CI] 3.1 ist im Gleichgewicht
 - Abbau Fibronektin und Bildung Katalysator (F`) [C]=[CA]+[CAF]+[CI] 3.1 ist im Gleichgewicht.")
35
Anwendung Massenwirkungsgesetz
Indirekter Inhibitor

36
Anwendung MM-Kinetik

37
Anwendung MM-Kinetik

38
Anwendung MM-Kinetik

39
Anwendung Massenwirkungsgesetz
c(x,t), C(x,y,t) – Konzentration Protelytisches Enzym ca(x,t), Ca(x,y,t) – Konzentration Aktive Protease ci(x,t), Ci(x,y,t) – Konzentration gehemmte Enzyme ia(x,t), Ia(x,y,t) – Konzentration Protease Inhibitor f(x,t), F(x,y,t) – Konzentration Fibronektin a(x,t), A(x,y,t) – Konzentration Angiostatin n(x,t), N(x,y,t) -EC Dichte v(x,t), V(x,y,t)- Konzentratin Angiogenetischer Faktor
, C(x,y,t) – Konzentration Protelytisches Enzym ca(x,t), Ca(x,y,t) – Konzentration Aktive Protease ci(x,t), Ci(x,y,t) – Konzentration gehemmte Enzyme ia(x,t), Ia(x,y,t) – Konzentration Protease Inhibitor f(x,t), F(x,y,t) – Konzentration Fibronektin a(x,t), A(x,y,t) – Konzentration Angiostatin n(x,t), N(x,y,t) -EC Dichte v(x,t), V(x,y,t)- Konzentratin Angiogenetischer Faktor")
40
Chemischer Transport in der Kapillaren

41
Chemischer Transport in der Kapillaren

42
Chemischer Transport in der ECM
Konzentrationsgradient beschreibt den Konzentrationsunterschied

43
Chemischer Transport in der ECM

44
Chemischer Transport in der ECM

45
Chemischer Transport in der ECM

46
Zellbewegung Bewegung der EC an der Kapillarwand:

47
Zellbewegung Θ: Proliferation (N-N0)/N0 Ca
/N0 Ca")
48
ECM-Kapillar Transmission
Verbindung ECM-Transport-Gleichung mit den Kapillar-Transport-Gleichungen:

49
Numerisch Simulation

50
Numerisch Simulation

51
Numerische Simulation

52
Numerische Simulation
~1-2mm vom Limbus(EC) entfernt Sprossung nach vier Tagen Vaskulär nach weniger als 7 Tagen, ca. 0,5mm Wachstum pro Tag Tumor mit 6mm Entfernung: avaskuläres Wachstum 0,1-0,2 mm pro Tag Kapillare wuchsen mit 1 mm pro Tag am Anfang
 entfernt. Sprossung nach vier Tagen. Vaskulär nach weniger als 7 Tagen, ca. 0,5mm Wachstum pro Tag. Tumor mit 6mm Entfernung: avaskuläres Wachstum 0,1-0,2 mm pro Tag. Kapillare wuchsen mit 1 mm pro Tag am Anfang.")
53
Numerische Simulation
Z-Achse: EC-Dichte Kapillarwachstum ohne Angiostatin (Distanz Tumor zu Kapillaren 25microns) Wachstumfaktor braucht 3,49h für Durchquerung der ECM
 Wachstumfaktor braucht 3,49h für Durchquerung der ECM.")
54
Numerische Simulation
Z-Achse: Fibronektin-Dichte Abbau von Fibronektin in der ECM, Bildung eines Tunnels

55
Numerische Simulation
Z-Achse: EC-Dichte EC-Ausbreitung nach der Gabe von Angiostatin (T=4,45h) Kein Abbau, aber wenig/keine Gefäßneubildung
 Kein Abbau, aber wenig/keine Gefäßneubildung.")
56
Eigenschaften Zellbewegung
Blow-up:

57
Eigenschaften Zellbewegung

58
Eigenschaften Zellbewegung

59
Eigenschaften Zellbewegung

60
Eigenschaften Zellbewegung

61
Eigenschaften Zellbewegung

62
Blow-up von P für N=2, D=0,04

63
Eigenschaften Zellbewegung
Definition: P(x,t) aggregiert, wenn es gegen einen nicht-konstanten stationären Zustand konvergiert für t endlich oder unendlich Massetransport entlang von Charakteristiken: ut+ux=0 hat die Lösung u(x,t)=f(x-t) und die Lösung ist entlang der Charakteristiken x-t=konstant
 aggregiert, wenn es gegen einen nicht-konstanten stationären Zustand konvergiert für t endlich oder unendlich. Massetransport entlang von Charakteristiken: ut+ux=0 hat die Lösung u(x,t)=f(x-t) und die Lösung ist entlang der Charakteristiken x-t=konstant.")
64
Eigenschaften Zellbewegung

66
w fixiert Neigungen der Charakteristiken in R1 haben selbes Vorzeichen, Massentransport in R2 E(x,t) Vorzeichenwechsel Masse in R3 bleibt dort (Grenze: E1(x,t))
)")
67
Für w0 groß und t klein

68
Für w0 klein und positiv, P(x,0)=1+εcos(2πx)
Nur zwei Regionen R0 und R1, „blow-up“ an Scheitelpunkt

69
Für w0 größer und positiv, treten regionen R2 und R3 auf
Nur zwei Regionen R0 und R1, „blow-up“ an Scheitelpunkt

73
Schocks können zu Aggregation führen
Ohne Dämpfunfgsterm können sich in t>0 Schocks entlang der Charakteristiken bilden: Charakteristiken konvex p´=u0´´(x)>0 Charakteristiken konkav p0´=u0´´<0 Schocks können zu Aggregation führen
>0. Charakteristiken konkav p0´=u0´´<0. Schocks können zu Aggregation führen.")
Ähnliche Präsentationen
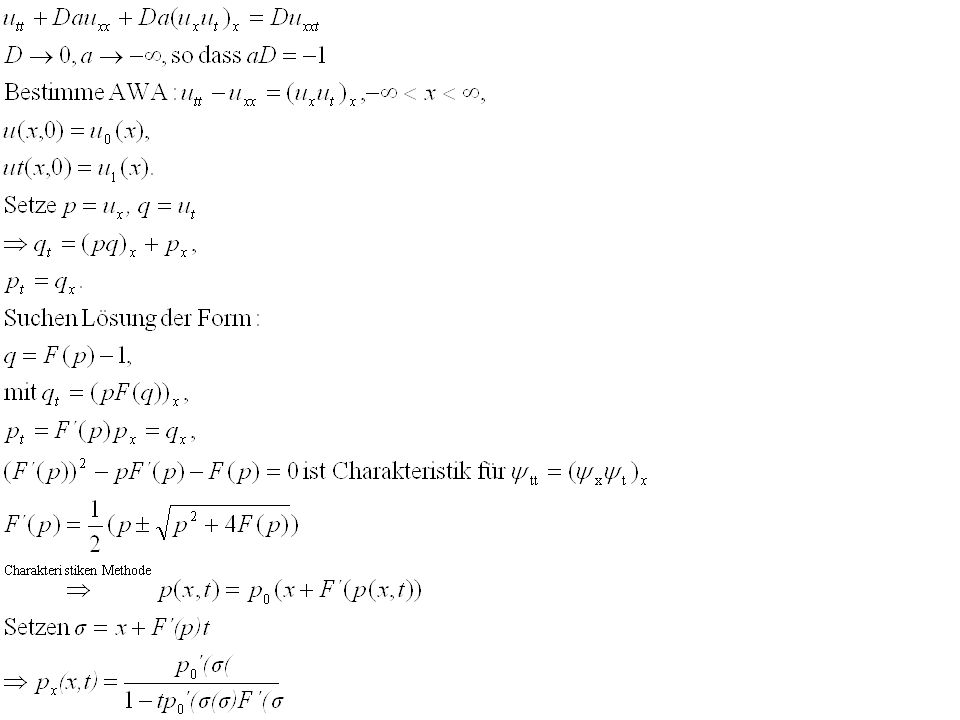















 Prof. Th. Ottmann.>")




>")

