Präsentation herunterladen
Die Präsentation wird geladen. Bitte warten

1
Non-conducting functions of voltage-gated ion channels
Leonard Kaczmarek Oktober 2006, nature review neuroscience

2
Einführung : Grundlagen für das Verständnis leitender Funktionen von spannungsabhängigen Ionenkanälen

3
Klassifizierung von Ionenkanälen
Gating Aktionspotential Alternatives Gating

4
Klassifizierung von Ionenkanälen
Kationenkanäle: hohe Selektivität (Namen!) Ionenkanäle Anionenkanäle: geringe Selektivität spannungsabhängig (präsynaptisch) Gating ligandenabhängig (postsynaptisch)
 Ionenkanäle. Anionenkanäle: geringe Selektivität. spannungsabhängig (präsynaptisch) Gating. ligandenabhängig (postsynaptisch)")
5
Gating 2 Zustände von Ionenkanälen: offen u. geschlossen
Energie für Konformationsänderung stammt bei den spannungsabh. Kanälen aus der Änderung der Membranspannung Kaskade von Konformationsänderungen

6
Aktivierung <-> Deaktivierung
Übertragung der Energie auf Spannungssensor des Kanals Bewegung des Sensors im elektrischen Feld Konsequenz: weitere Konformationsänderungen im Kanal Resultat: Öffnung der Pore In Nav-Kanälen geschieht dies in weniger als einer Millisekunde, bei Kv-Kanälen dauert es deutlich länger: 10 bis ungefähr 70 ms.

7
Kanalinaktivierung (N-Typ)
bei Nav-Kanälen und bei einigen Kv-Kanälen (A-Typ-Kanäle) führt zur Unterbrechung des Ionenstroms Innerhalb weniger Millisekunden nach Aktivierung
 führt zur Unterbrechung des Ionenstroms. Innerhalb weniger Millisekunden nach Aktivierung.")
8
Weitere Mechanismen C-Typ-Inaktivierung:
Beruht auf Konformationsänderungen im Bereich des Selektivitätsfilters unabh. aber beschleunigt durch N-Typ-Inaktivierung besonders langsame Rückreaktion

9
Ruhepotential Nahe beim Gleichgewichtspotential für Kalium: -70 mV
Leitfähigkeit für Kalium über Kir-Kanäle oder 2-P-Domänen Kaliumkanäle sichergestellt

10
Aktionspotential I: Initiationsphase II: Depolarisation IIa: Aufstrich IIb: Overshoot III: Repolarisation IV: Nachhyperpolarisation Transiente Änderung des Membranpotentials, ausgelöst durch einen Reiz, der die Zelle über einen Schwellenwert hinaus depolarisiert.

11
Depolarisation Stimulus depolarisiert Membranpotential bis zum Schwellenwert Spannungsabhängige Natriumkanäle (Nav) öffnen sich, Na+ strömt in die Zelle Kir-Kanäle (o. Ä.) werden inaktiviert
 öffnen sich, Na+ strömt in die Zelle. Kir-Kanäle (o. Ä.) werden inaktiviert.")
12
Repolarisation Inaktivierung der Nav-Kanäle Öffnen der Kv-Kanäle

13
Nachhyperpolarisation
Ca2+-abhängige Kaliumkanäle Konzentration wird mit der Zeit gesenkt

14
alternatives Gating Ionenkanäle können durch versch. Signale geöffnet bzw. verschlossen werden: intrazelluläre Messenger Proteine mechanische Spannung Wärme/Kälte kleinmolekulare Porenblocker

15
Intrazelluläre Messenger
ATP an 2-Segment-Kalium-Kanälen (A) cAMP und cGMP an HCN- und CNG-Kanälen (B) Andere über: pH, oder Ca2+
 cAMP und cGMP an HCN- und CNG-Kanälen (B) Andere über: pH, oder Ca2+")
16
HCN-Kanäle nicht-selektive Kationenkanäle verantwortlich für Ih-Strom
Langzeitbahnung der synapt. Antworten, Ausschüttung von Neurotransmittern über mehrere Stunden Initiieren Langzeitbahnung

17
physikalische Faktoren
Umgebungsqualitäten (Kälte, Wäreme, mechanische Zugkraft und Osmolarität) können in Kanalgating umgesetzt werden. -> hierfür verantwortlich: TRP-Kanäle
 können in Kanalgating umgesetzt werden. -> hierfür verantwortlich: TRP-Kanäle.")
18
Nichtleitende Funktionen spannungsabhängiger Ionenkanäle

19
Gliederung - Einführung: altes und neues Verständnis - Kaliumkanäle - Natriumkanäle - Calciumkanäle - TRP – Kanäle - Nichtleitende Funktionen von Ionenkanälen und Krankheiten - Fazit

20
Einführung: altes und neues Verständnis
- primäre Funktion spannungsabhängiger Ionenkanäle: Erzeugung verschiedener Typen von Feuermustern für Informationsprozessierung und motorische Outputs - Modulierbarkeit (enzymatisch) ermöglicht Anpassung elek- trischer Eigenschaften des Neurons an die Inputsituation Diese Modulation wird üblicherweise als einseitiger Prozess betrachtet, bei welchem die Ionenkanäle als passive Elemente verändert werden
 ermöglicht Anpassung elek- trischer Eigenschaften des Neurons an die Inputsituation. Diese Modulation wird üblicherweise als einseitiger Prozess betrachtet, bei welchem die Ionenkanäle als passive Elemente verändert werden.")
21
- aktuell (bis Okt. 2006) : Hinweise auf direkte Beeinflussung biochemischer Ereignisse in Zellen durch Ionenkanäle ohne (direkte) Notwendigkeit der leitenden Funktion als Ionenkanal - direkte Aktivierung von Enzymen intrazellulärer Signalwege - Funktion als Adhäsionsmoleküle oder Komponenten des Cytoskelettes - Regulation der Expression bestimmter Gene
 Notwendigkeit der leitenden Funktion als Ionenkanal. - direkte Aktivierung von Enzymen intrazellulärer Signalwege - Funktion als Adhäsionsmoleküle oder Komponenten des Cytoskelettes - Regulation der Expression bestimmter Gene.")
22
methodische Grundlagen und Ergebnisse zitierter Studien :
- Deletion (/Knockout) / Überexpression / Transfektion verändert Zelleigenschaften auf Arten, die nicht durch bloße Veränderung der Erregbarkeit erklärt werden können - Untersuchungen mit nichtleitenden Mutanten gleiche Effekte - vom Ionenfluss unabhängige Funktionen bestimmter Untereinheiten gefunden
 / Überexpression / Transfektion verändert Zelleigenschaften auf Arten, die nicht durch bloße Veränderung der Erregbarkeit erklärt werden können - Untersuchungen mit nichtleitenden Mutanten gleiche Effekte. - vom Ionenfluss unabhängige Funktionen bestimmter Untereinheiten gefunden.")
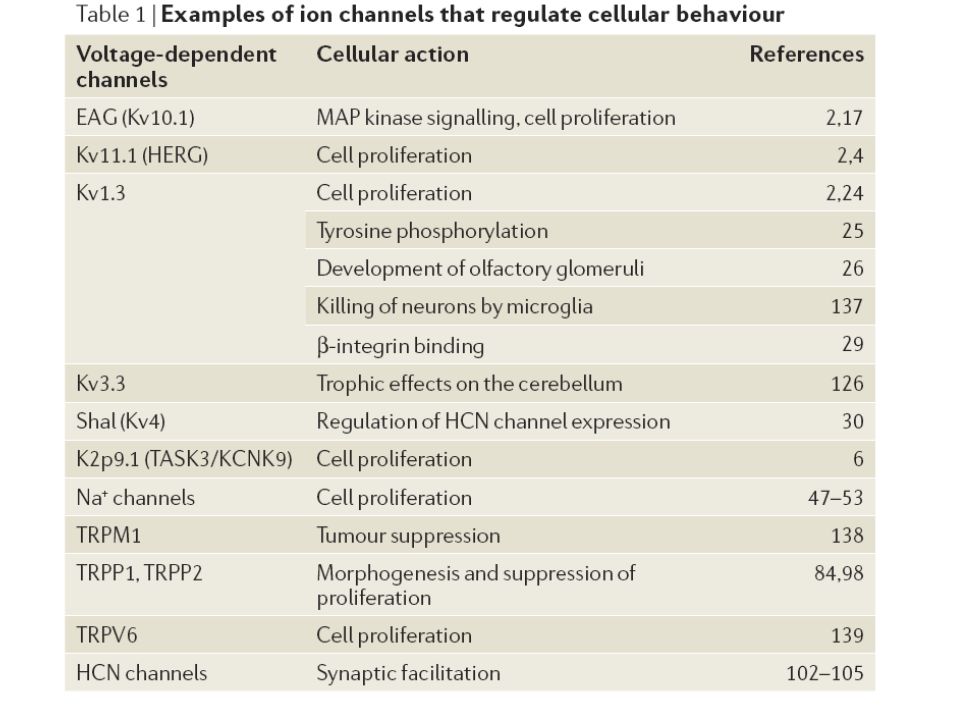
25
Kaliumkanäle erhöhte Expression / Aktivität in Tumorzellen Hinweis auf Zell-regulative Funktionen - Transfektion in Gliazellen / nicht- neuronale Zellen : Initiierung / Beschleunigung der Zellproliferation These: direkter Einfluss auf cytoplasmatische und nukleare Signalwege

26
Kv10.1 K+ - Kanal (EAG) - bisher als weitgehend typisch für synaptische Enden von Neuronen betrachtet - Drosophila melanogaster mit eag – knockout : Habituation und assoziatives Lernen verändert

27
Besondere Eigenschaften der cytoplasmatischen Domänen:
- Drosophila: EAG bindet eine Kinase (CaMKII; α- UE), was zu deren konstitutiver Aktivierung führt - (C)- Terminus : nuclear localization signal - (N) – Terminus: PAS – Domäne (bei anderen Proteinen regulativer Einfluss auf Transkription und Proteinkinase Aktivität)
, was zu deren konstitutiver Aktivierung führt. - (C)- Terminus : nuclear localization signal - (N) – Terminus: PAS – Domäne (bei anderen Proteinen regulativer Einfluss auf Transkription und Proteinkinase Aktivität)")
28
Beteiligung des Spannungssensors an regulativen Einflüssen:
- Beeinflussung der Aktivität des Signalweges der Mitogen-aktivierten Proteinkinase (MAPK) - Transfektionsexperimente mit Fibro- / Myoblasten: Erhöhung von Zelldichte und DNA –Synthese unter gesteigerter Aktivität der p38 MAPK; unabhängig von der Leitfähigkeit des EAG, aber abhängig von Gating – Zustand des Kanals !
 - Transfektionsexperimente mit Fibro- / Myoblasten: Erhöhung von Zelldichte und DNA –Synthese unter gesteigerter Aktivität der p38 MAPK; unabhängig von der Leitfähigkeit des EAG, aber abhängig von Gating – Zustand des Kanals !")
29
Einfluss des Gatingzustandes auf regulative Funktionen des EAG - Kanals
- Verwendung von Mutanten, bei welchen die Spannungsab- hängigkeit der Aktivierung in den Bereich negativer Potentiale verlagert ist Kanal dauerhaft geöffnet - Folge : Blockade des proliferativen Effekts EAG Kanal muss sich zur Aktivierung der p38MAPK im geschlossenen Zustand befinden [Beleg für die Unabhängigkeit des Effekts vom Ionenfluss !]

30
Kv1.3 - Kanal - u.a. im Hippocampus und bulbus olfactorius exprimiert
- besondere Eigenschaft: zeigt bei wiederholter Depolarisation kumulative Inaktivierung (K+ - Kanäle: Repolarisation ) zeitliche Ausdehnung von AP; erhöhte Erregbarkeit bei wiederholter Stimulation des Neurons
 zeitliche Ausdehnung von AP; erhöhte Erregbarkeit bei wiederholter Stimulation des Neurons.")
31
Effekte der Aktivität des KV1.3 – Kanals auf Zellproliferation
-in Transfektionsexperimenten vom Ionenstrom durch den Kanal abhängig gewesen ! (Porenmutanten /K+-Ionophore) In vivo – Hinweise : neben Beeinflussung des Membranpo tentials und K+ - Strom weitere Aspekte relevant
 In vivo – Hinweise : neben Beeinflussung des Membranpo- tentials und K+ - Strom weitere Aspekte relevant.")
32
Experimente mit Kv1.3 knockout - Mäusen
- deutlich erhöhte Level von Signalmolekülen im bulbus olfactorius - niedrigere Konzentrationsschwelle für das Entdecken von Duftstoffen - erhöhte Fähigkeit zur Unterscheidung verschiedener Duftstoffe - diese Effekte traten nicht beim Eliminieren anderer K+ - Kanäle auf Ursache nicht auf bloße Veränderung der Membran- erregbarkeit beschränkt

33
möglicher Mechanismus des spannungsunabhängigen Effektes :
- Kv1.3 in intakten Zellen eng mit β – Integrinen assoziiert - diese regulieren Zell –Zell- Interaktionen, Zellwachstum und -überleben - spezifische Kv1.3 – Blocker heben die Assoziation mit Integrinen auf Kv1.3 – Aktivität trägt vermutlich direkt zur Integrin – Regulation bei

34
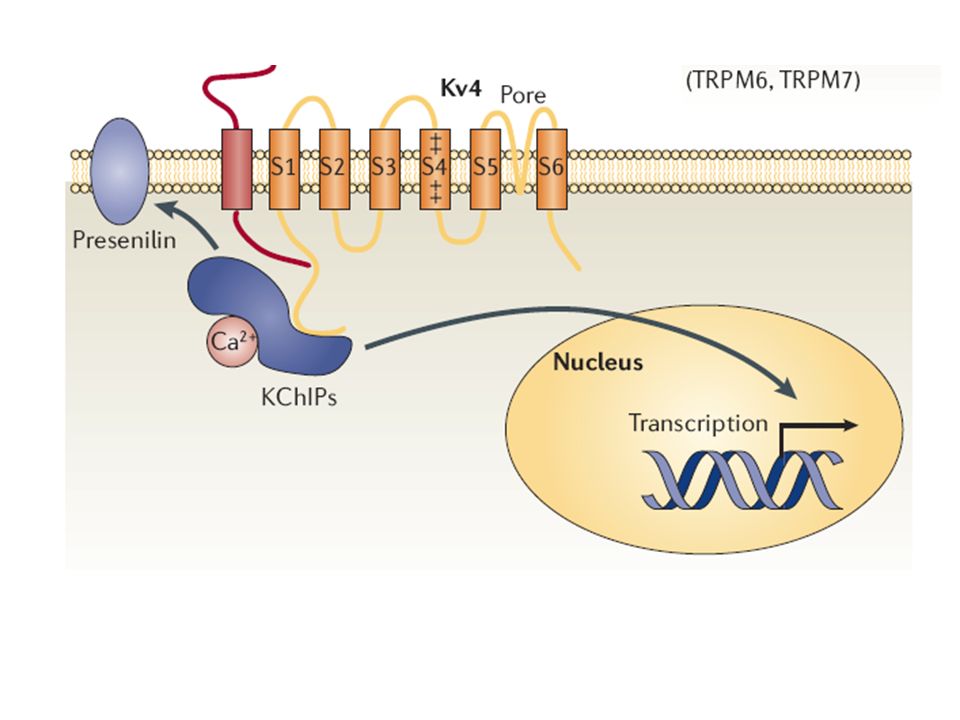
Koordination neuronaler Erregbarkeit
- Koordination der Expression von Genen für Kanäle bis dato ungeklärt aktuelle Studien : - Expressionslevel von Kv4 – Kanälen beeinflusst Expression eines anderen Kanals; dieser Effekt könnte vom Ionenfluss unabhängig sein - genauer Mechanismus noch unklar

35
Enzymatische Aktivität der β – Untereinheiten von Kv - Kanälen
- neben Assoziation von Kinasen (EAG;α- UE) häufig Bindung zusätzlicher Untereinheiten gefunden - einige dieser zusätzlichen UE zeigen enzymatische Aktivität - möglicherweise vermitteln sie einige der Effekte von K Kanälen auf zelluläre Signalwege
 häufig Bindung zusätzlicher Untereinheiten gefunden. - einige dieser zusätzlichen UE zeigen enzymatische Aktivität. - möglicherweise vermitteln sie einige der Effekte von K+ - Kanälen auf zelluläre Signalwege.")
36
Bsp.: Familie des K+ - Channel interacting protein (KChIP1-4)
- direkte Bindung an Kv4 α – UE, welche schnell inaktivierende A-Typ Kaliumauswärtsströme erzeugen Beeinflussung von Amplitude und Kinetik der Kv4 – Inaktivierung durch KChIP1-4 - ferner Funktion als Ca2+ -bindende Regulatoren von Presenilinen (membranintegrale Spaltenzyme) und Ca2+ - abhängige Transkriptionsfaktoren
 und Ca2+ - abhängige Transkriptionsfaktoren.")
38
Zusammenfassung: nichtleitende Funktionen von K+ - Kanälen
- direkter Einfluss auf cytoplasmatische und nukleare Signalwege - Effekte auf Signalübertragung können z.B. vom Gatingzustand abhängen - Effekte beeinflussen perzeptuelle / kognitive Fähigkeiten durch Regulation verschiedener zellulärer Prozesse und können darüber hinaus Ausbruch von Krankheiten begünstigen

39
Na+ - Kanäle

40
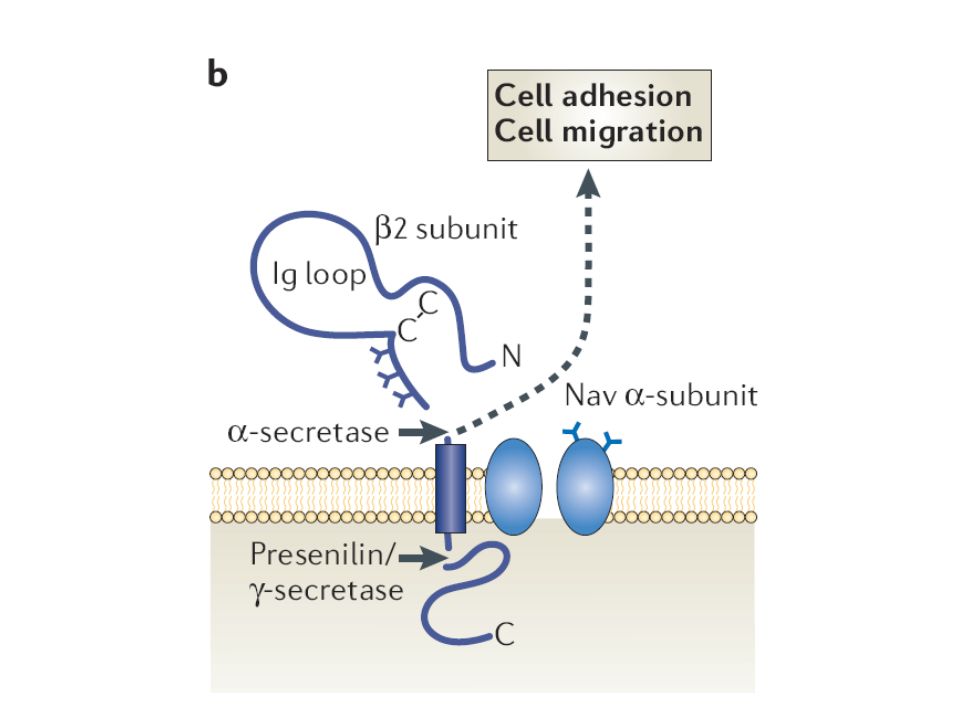
Na+ - Kanal β – Untereinheiten als Adhäsionsmoleküle
- CAMs (Cell Adhesion Molecules) mit kurzem cytoplasma- tischem C – Terminus und langen extrazellulären Domänen vermitteln Zell – Zell – Adhäsion durch homo- oder hetero- typische Interaktionen und Interaktionen mit Molekülen der extrazellulären Matrix
 mit kurzem cytoplasma- tischem C – Terminus und langen extrazellulären Domänen. vermitteln Zell – Zell – Adhäsion durch homo- oder hetero- typische Interaktionen und Interaktionen mit Molekülen der extrazellulären Matrix.")
41
- an der cytoplasmatischen Seite Bindung an Proteine, welche ihrerseits mit dem Cytoskelett verbunden sind (Ankyrin G undB) β – Untereinheiten ermöglichen Na+ - Kanälen eine physikalische Verbindung des Cytoskeletts mit der Oberfläche von Nachbarzellen - vermutlich trägt diese Interaktion zur Ausbildung von Axonhügeln und Ranvier´schen Schnürringen bei (hohe Dichte von Na+ - Kanälen)
")
43
Einflüsse der Untereinheiten auf das Wachstum von Neuriten
- β1 – Untereinheiten können das Wachstum von Neuriten bei dissoziierten Neuronen stimulieren - β2 – Untereinheiten dagegen können die Länge von Neuriten durch einen Effekt reduzieren, der durch α- oder β1- Unter- einheiten aufgehoben werden kann nicht- leitende Funktionen von Na+ - Kanälen könnten wichtige organisatorische Elemente der neuronalen Entwicklung sein

44
Regulation von β –Untereinheiten der Na+ - Kanäle via Spaltung
- β –Untereinheiten werden kontinuierlich durch proteolytische Spaltung umgesetzt - möglicherweise reguliert Spaltung der extrazellulären und anschließend der (C) – terminalen Domäne die Beteiligung der β2 – Untereinheit an Prozessen der Zelladhäsion und -migration
 – terminalen Domäne die Beteiligung der β2 – Untereinheit an Prozessen der Zelladhäsion und -migration.")
46
Ca2+-Kanäle Ca2+-Kanäle wohl am engsten verschränkt mit zellulären Ereignissen Wie kann es sein, dass versch. Wege der Konzentrationserhöhung von Ca2+, verschiedene Effekte auf die Zelleigenschaften haben? Beispiel: Aktivierung des Ca2+-Einstroms durch L-Typ Ca2+-Kanäle der Cav-Familie, aktiviert die MAPK-ERK-Signalkaskade. Dadurch wird CREB, ein Transkriptions-faktor aktiviert.

47
elektromech. Kopplung in Skelettmuskelzellen
Signal unabhängig vom Ionenfluss phys. Verbindung des Ca2+-Kanals mit Ryanodin-Rezeptor -> Reaktion als Kanal, aber auch als Spannungssensor für Ryanodin-Rezeptor Bei der elektromechanischen Kopplung in Skelettmuskelzellen erzeugt die Aktivierung eines Ionenkanals ein Signal, welches unabhängig ist von der Fähigkeit des Ionenflusses. Hier führt eine Depolarisation zu einer Aktivierung von (L-Typ) spannungsabhängigen Calcium-1.1-Kanälen (Cav). Aber auch bei Abwesenheit von externem Ca2+ führt die Konformationsänderung im Kanal, die durch die Depolarisation hervorgerufen wird zu einer Muskelkontraktion. Die cytoplasmatische Domäne des Kanals ist mit dem Ryanodin-Rezeptor verbunden, über dessen Kanal aus dem sarkoplasmatischen Retikulum Ca2+ freigegeben werden kann. Cav1.1-Proteine agieren also als normale Ca2+-Kanäle, aber eben auch als Spannungssensoren, welche direkt eine Konformationsänderung im Ryanodin-Rezeptor/Cav1.1-Komplex hervorrufen, wodurch dann Ca2+ aus dem SR freigegeben werden kann.
 spannungsabhängigen Calcium-1.1-Kanälen (Cav). Aber auch bei Abwesenheit von externem Ca2+ führt die Konformationsänderung im Kanal, die durch die Depolarisation hervorgerufen wird zu einer Muskelkontraktion. Die cytoplasmatische Domäne des Kanals ist mit dem Ryanodin-Rezeptor verbunden, über dessen Kanal aus dem sarkoplasmatischen Retikulum Ca2+ freigegeben werden kann. Cav1.1-Proteine agieren also als normale Ca2+-Kanäle, aber eben auch als Spannungssensoren, welche direkt eine Konformationsänderung im Ryanodin-Rezeptor/Cav1.1-Komplex hervorrufen, wodurch dann Ca2+ aus dem SR freigegeben werden kann.")
48
außerdem: Verbindung zwischen Ca2+-Kanälen und dem Exocytose-Komplex
Interaktion mit SNARE-Proteinen -> enge Verbindung zwischen Vesikeln und den Kanälen

49
Hinweise, dass die physikalische Verbindung Informationen über den Zustand der Kanäle zur release site übertragen kann Ausschüttung von Neurotransmittern unabhängig vom Ca2+-Einstrom Depolarisation der Membran führt zur Konformationsänderung im Kanal und einer Änderung der Aktivität der SNARE-Proteine Effekt kann verhindert werden durch intrazelluläre Blockade der Bindung zwischen Cav2.1-Kanälen und den SNARE-Proteinen

50
TRP-Kanäle nicht-selektive Kationenkanäle
Einstellung des Ruhemembranpotentials, regulieren Konzentrationen von Ca2+ und Mg2+ in Neuronen und anderen Zellen Überexpression in Krebszellen

51
TRP-Kanäle sind eingeteilt in sechs Unterfamilien
TRPP1 & 2 unterdrücken Zellproliferation In TRPM gibt es längere cytoplasmat. C-terminale Domänen als in den anderen Unterfamilien Proteinkinaseaktivität bei TRPM6 & 7 (einzige Kanäle mit enzymatischer Aktivität in einer porenbildenden a-UE)
")
52
Nichtleitende Funktionen von Ionenkanälen und Krankheiten
- Mutationen an Genen bestimmter Ionenkanäle können über Veränderung der nichtleitenen Funktionen zu Veränderungen der Intrazellulären Signalwege… führen diese führen schließlich zu Veränderungen in der Organisation des Nervensystems - Beispiele: Chorea Huntington (Na+ - Kanäle), Schizophrenie (K+ - Kanäle)
, Schizophrenie (K+ - Kanäle)")
53
Fazit - Verständnis von nichtleitenden Funktionen spannungsab- hängiger Ionenkanäle und deren Bedeutung immer noch begrenzt - trotz einiger klarer Beispiele insbesondere Kopplung an Signalwege noch unklar - konzeptionelles Problem: In vielen Fällen könnte ein Ionenfluss für den biotischen Effekt nötig sein

54
- Funktionen nichtleitender Eigenschaften von Ionenkanälen könnten die Vielzahl an Genen (beachte ferner posttran- skriptionale Modifikationen) für Ionenkanalproteine, die Vielzahl von Kanälen mit geringen elektrischen Eigen- schaften und das hohe Maß an Konservierung der Gene erklären
 für Ionenkanalproteine, die Vielzahl von Kanälen mit geringen elektrischen Eigen- schaften und das hohe Maß an Konservierung der Gene erklären")
Ähnliche Präsentationen







>")





 und der cytoplasmatischen Tyrosinkinase c-src in Urothelkarzinomzellen A. Melchior, J. Herrmann,>")







