Präsentation herunterladen
Die Präsentation wird geladen. Bitte warten

1
Zur Erinnerung... -Der Einschluss der DNA in ein eigenes Kompartiment, dem Zellkern, machte einen intensiven Stoffaustausch zwischen Cytoplasma und Kern notwendig. -Ort des Transports ist die Kernpore, eine der größten Strukturen in der Zelle (125MDa, ca Proteine). -Kleine Proteine und Metaboliten können passiv in den Kern gelangen. Große Moleküle (40kDa und größer) benötigen hingegen einen aktiven und Rezeptor-vermittelten Transportprozess. -Moleküle die in den Kern transportiert werden haben ein sogenanntes nuclear location signal (NLS, das klassische Signal ist PKKKRKVE). -Importrezeptoren sind die Importine. Für die klassische NLS gibt es ein Importin-heterodimer das man Importin a/ß nennt. -Die Importine können das Cargo nur an die Kernpore bringen (docking). Für die nachfolgende Translokation ist der Transportfaktor Ran und GTP notwendig.
. -Kleine Proteine und Metaboliten können passiv in den Kern gelangen. Große Moleküle (40kDa und größer) benötigen hingegen einen aktiven. und Rezeptor-vermittelten Transportprozess. -Moleküle die in den Kern transportiert werden haben ein sogenanntes. nuclear location signal (NLS, das klassische Signal ist PKKKRKVE). -Importrezeptoren sind die Importine. Für die klassische NLS gibt es ein. Importin-heterodimer das man Importin a/ß nennt. -Die Importine können das Cargo nur an die Kernpore bringen (docking). Für die nachfolgende Translokation ist der Transportfaktor Ran und GTP. notwendig.")
2
Das Importin a/ß Dimer ist nicht für den Transport ausreichend
Docken ist möglich, nicht aber die Translokation in den Zellkern

3
Eine kleine GTPase reguliert den Kerntransport

4
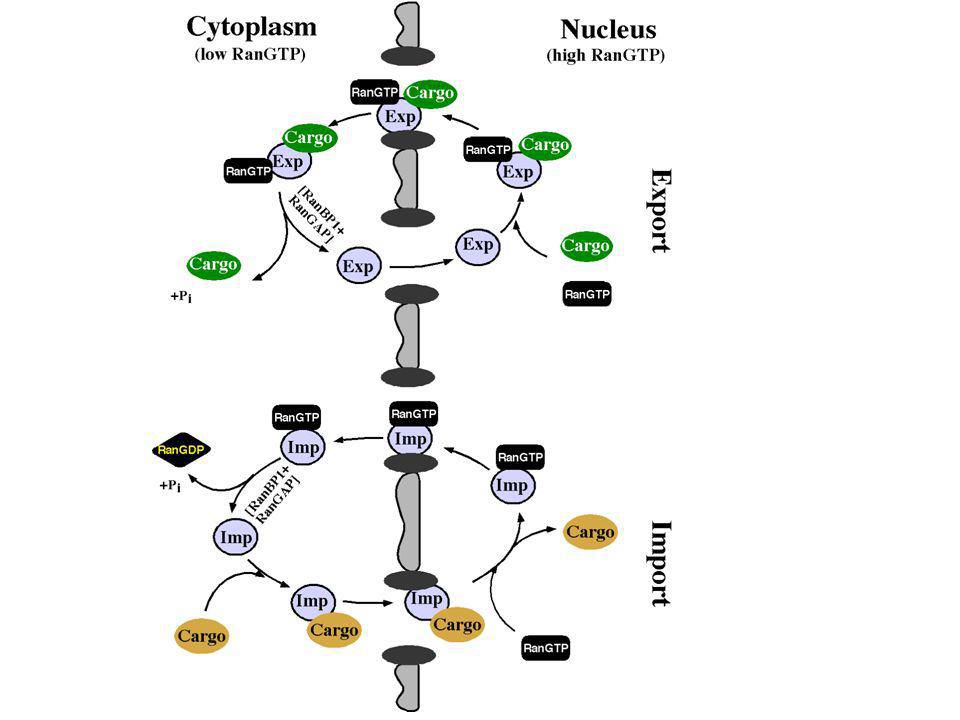
Die Rolle von Ran beim Kernimport
RanGTP dissoziiert den Importin a/b Komplex

5
RanGDP RanGTP RanGDP RanGTP

7
Der Ran-Zyklus bestimmt die Richtung des Transportvorgangs

8
Der Kernexport von Cargo
-viele Komponenten des Zellkerns müssen auch exportiert werden (mRNAs, Ribosomen, diverse Proteine etc.) Der Export ist ebenfalls durch spezifische Signale, den Nuclear Export Signals (NES) vermittelt. Diese Signale liegen i.d.R. auf spezifischen Proteinen LPP LER LTL die klassische NES vom HIV1 Rev Leucin-Reste sind sehr wichtig (hydrophob, vgl. NLS)
 Der Export ist ebenfalls durch spezifische Signale, den. Nuclear Export Signals (NES) vermittelt. Diese Signale liegen. i.d.R. auf spezifischen Proteinen. LPP LER LTL die klassische NES vom HIV1 Rev. Leucin-Reste sind sehr wichtig. (hydrophob, vgl. NLS)")
9
Der Kernexport von Cargo
- Die NES wird im Zellkern von spezifischen Exportrezeptoren, den Exportinen erkannt. -Im Gegensatz zum Import ist die Interaktion von NES mit Exportin aber nur in Anwesenheit von RanGTP möglich. Ran GDP dissoziiert hingegen den NES-Exportin-Komplex RanGDP

11
Zurück zur Tagesordnung: Der Abbau von Proteinen
und Aminosäuren -Der Abbau von Proteinen ist Quelle von Aminosäuren, die an mehreren Stellen im Metabolismus Verwendung finden. Der Abbau von Proteinen findet durch die Verdauung von Nahrung und durch die gezielte Degradation über das Ubiquitin-System in der Zelle statt. - Aminosäure sind Bausteine für Proteine und Vorstufen für andere (häufig stickstoffhaltige) Verbindung wie z.B. Nucleotidbasen. Amino- säuren können nicht als Energiespeicher dienen, sondern werden direkt in den Energiestoffwechsel eingeschleust. -Die Einschleusung der meisten Aminosäuren in den Energie- stoffwechsel erfolgt über den Harnstoff-Zyklus.
 Verbindung wie z.B. Nucleotidbasen. Amino- säuren können nicht als Energiespeicher dienen, sondern werden direkt. in den Energiestoffwechsel eingeschleust. -Die Einschleusung der meisten Aminosäuren in den Energie- stoffwechsel erfolgt über den Harnstoff-Zyklus.")
12
Die Aufnahme von Aminosäuren durch Nahrung
Zymogene des Pankreas Chymotrypsinogen Trypsinogen Procarboxypetidase Proelastase spezifische Transporter Pepsinogen (pH 2-Optimum) (Magen)
 (Magen)")
13
Die Ubiquitin-abhängige Degradation von Proteinen
- Jedes Protein hat eine charakteristische Halbwertzeit (von wenigen Minuten bis Jahren). -Defekte Proteine müssen ebenfalls abgebaut werden. -Proteine, die abgebaut werden sollen, werden mit einem spezifischen „Marker“ versehen. Dieser Marker ist ein kleines Protein („Ubiquitin“), das kovalent an das zu degradierende Protein genüpft wird. -Ubiquitin ist hochkonserviert und bei allen Eukaryonten zu finden (Unterschied zwischen Hefe und Mensch: 3 As von 76)
. -Defekte Proteine müssen ebenfalls abgebaut werden. -Proteine, die abgebaut werden sollen, werden mit einem spezifischen. „Marker versehen. Dieser Marker ist ein kleines Protein („Ubiquitin ), das kovalent an das zu degradierende Protein genüpft wird. -Ubiquitin ist hochkonserviert und bei allen Eukaryonten zu finden. (Unterschied zwischen Hefe und Mensch: 3 As von 76)")
14
Markierung mit Ubiquitin
C-Terminus von Ubiquitin bindet an ein oder mehrere Lysin-Reste (=verzweigtes Protein)
")
15
Tetraubiquitin: Das Abbausignal
-Lysin 48 eines Ub ist mit C-Terminus eines zweiten Ubs verküpft

16
Die Ubiquitinkonjugation
E1: aktivierendes Enzym. Bildet Adenylat-Zwischenprodukt und Thioester zwischen Ub und E1 (vlg. Aktivierung von FS in ß-Oxid.) E2: konjugierendes Enzym (Übernimmt aktiviertes Ub von E1) E3: katalysiert den Transfer auf das Zielprotein (Spezifität!)
 E2: konjugierendes Enzym (Übernimmt aktiviertes Ub von E1) E3: katalysiert den Transfer auf das Zielprotein (Spezifität!)")
17
Die Spezifität der Ubiquitinylierung
-Es gibt nur sehr wenige E1 Enzyme. -Es gibt eine größere Anzahl von E2 Enzymen, die untereinander homolog sind. -Es gibt viele E3-Enzyme, die zu unterschiedlichen Gruppen gehören. Die Kombination von E2 mit E3-Enzymen bei der Übertragung von Ub ermöglicht ein sehr großes Substratspektrum.

18
Wann wird ein Protein degradiert?
-bei Defekten (Missfaltungen, Oxidationen, Hydrolysen etc.). -der N-Terminus bestimmt die Halbwertszeit eines „normalen“ Proteins (N-end rule, N-Terminus Regel). -Spezifische Sequenzen im Protein können dieses destabilisieren (PEST- und cyclin destruction box-Sequenzen)
. -der N-Terminus bestimmt die. Halbwertszeit eines „normalen Proteins (N-end rule, N-Terminus. Regel). -Spezifische Sequenzen im Protein. können dieses destabilisieren (PEST- und cyclin destruction box-Sequenzen)")
19
Das Proteasom degradiert Ub-markierte Proteine
Bindet Ub-konjugiertes Substrat Isopeptidase spaltet Ub vom Protein regulatorische Einheiten Unter ATP verbrauch wird Substrat entfaltet katalytische Einheit Proteasom enthält Serin und Threonin- Peptidasen.

20
Fehlerhaftes zerstören von Proteinen kann Krankheiten hervorrufen
-Humanes Papillomavirus (HPV) codiert für eine spezifische E3-Ligase. Diese Ligase zerstört den Tumor-Supressor p53 (ein Protein, welches u.a. die DNA-Reparatur kontrolliert). Die Virus-induzierte Degradation von p53 führt häufig zu Krebs, da u.a. die DNA nicht mehr angemessen repariert werden kann!
 codiert für eine spezifische. E3-Ligase. Diese Ligase zerstört den Tumor-Supressor p53 (ein Protein, welches. u.a. die DNA-Reparatur kontrolliert). Die Virus-induzierte Degradation von p53 führt häufig zu Krebs, da u.a. die DNA nicht mehr angemessen repariert werden kann!")
21
Der Abbau der Aminosäuren
-viele Aminosäuren transferieren die Aminogruppe auf a-Ketoglutarat wobei Glutamat entsteht. Glutamat kann dann desaminiert werden.

22
Die Transaminase-Reaktion
-Der Transfer der a-Aminogruppe auf eine a-Ketosäure wird durch Aminotransferasen (auch Transaminase genannt) katalysiert. Bsp: Aspartat-Aminotransferase (OAA) Alanin-Aminotransferase (Pyruvat) Merke: Diese Reaktionen sind reversibel, und können so auch für die Synthese von Aminosäuren aus a-Ketosäuren verwendet werden!
 katalysiert. Bsp: Aspartat-Aminotransferase (OAA) Alanin-Aminotransferase (Pyruvat) Merke: Diese Reaktionen sind reversibel, und können so auch für. die Synthese von Aminosäuren aus a-Ketosäuren verwendet werden!")
23
Die Transaminase-Reaktion
-Der Transfer der a-Aminogruppe auf eine a-Ketosäure wird durch Aminotransferasen (auch Transaminase genannt) katalysiert. Bsp: Aspartat-Aminotransferase (OAA) Alanin-Aminotransferase (Pyruvat) Merke: Diese Reaktionen sind reversibel, und können so auch für die Synthese von Aminosäuren aus a-Ketosäuren verwendet werden!
 katalysiert. Bsp: Aspartat-Aminotransferase (OAA) Alanin-Aminotransferase (Pyruvat) Merke: Diese Reaktionen sind reversibel, und können so auch für. die Synthese von Aminosäuren aus a-Ketosäuren verwendet werden!")
24
Der Kofaktor aller Transaminasen Hydroxylgruppe die leicht sauer ist.
Pyridingring, der leicht basisch ist.

25
Der Kofaktor Pyridoxalphosphat
die Aldehydgruppe ist wichtig für die Transaminierungsreaktion, da sie Schiff-Basen mit primären Aminen ausbilden kann

26
As aus dem aktiven Zentrum
Ein internes Aldimin wird zu einem externen Aldimin As aus dem aktiven Zentrum des Enzyms As die desaminiert werden soll

27
Der Mechanismus der Transaminierung: 1. Schritt
Eine Elektronenfalle

28
Der Mechanismus der Transaminierung: 2. Schritt

29
Der Mechanismus der Transaminierung: 3. Schritt

30
Der zweite Teilschritt der Transaminierung
Der zweite Teilschritt ist die Umkehrreaktion der ersten z.B. kann a-Ketoglutarat jetzt mit dem PMP zum Glutamat reagieren.

31
Die oxidative Desaminierung: 1.Schritt
Glutamat-Dehydrogenase Vergleiche zur Oxidation von Alkoholen

32
Die oxidative Desaminierung: 1.Schritt
-Die Hydrolyse der Schiff-Base setzt das Ammoniak frei. -a-Ketoglutarat kann dann an einer neuen oxidativen Desaminierung teilnehmen.

33
Die Bilanz der oxidativen Desaminierung

34
Die Regulation der Glutamat-DH
-Glutamat ist ein allosterisches Enzym mit 6 UE -GTP und ATP sind allosterische Inhibitoren. -GDP und ADP sind allosterische Aktivatoren. -Die Erniedrigung der Energieladung der Zelle beschleunigt den Abbau von Aminosäuren.

Ähnliche Präsentationen






>")
>")












