Präsentation herunterladen
Die Präsentation wird geladen. Bitte warten

3
Überblick: Kommunikationsbahnen
Die Kegelschnecke tötet ihre Beute durch Gift, das die Neurone außer Gefecht setzt Neurone sind Nervenzellen, die Information innerhalb des Körpers weiterleiten Die Kommunikation über Neurone erfolgt weitgehend über zwei unterschiedliche Signaltypen: elektrische Signale, die über weite Strecken fortgeleitet werden, und chemische Signale, die auf Kurzstrecken eingesetzt werden

5
Die Informationen werden in Form elektrischer Signale weitergeleitet, die von Ionenbewegungen getragen werden Die Signalverarbeitung höherer Ordnung wird weitgehend von Neuronengruppen übernommen, die zu einem Gehirn organisiert sind oder aber einfachere Zellgruppen bilden, die als Ganglien bezeichnet werden

6
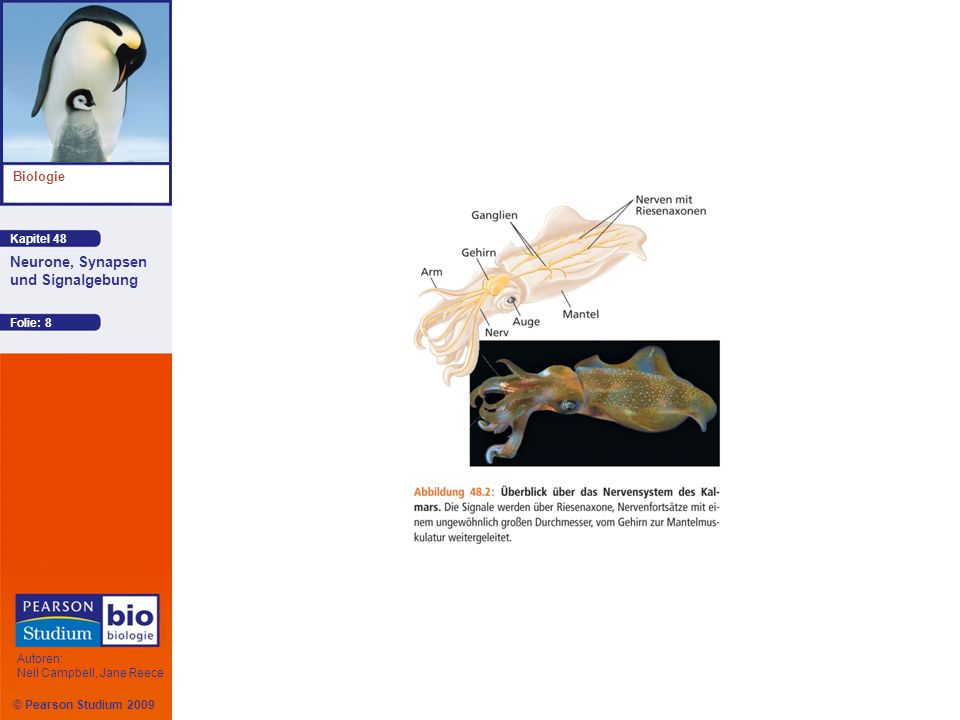
48.1: Neuronale Organisation und Struktur als Spiegel der Funktion bei der Informationsübermittlung
Der Kalmar ist ein Kopffüßer mit außerordentlich großen Nervenzellen, weshalb er sich bestens für Untersuchungen der Nervenfunktionen eignet

7
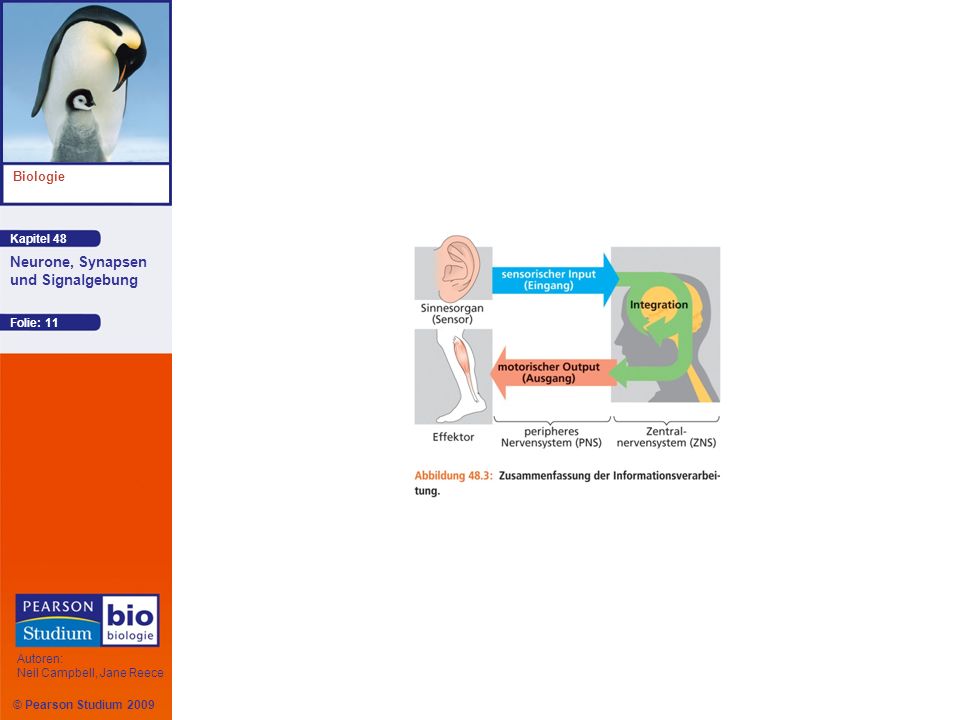
Einführung in die Informationsverarbeitung
Nervensysteme durchlaufen drei Stadien der Informationsverarbeitung: Verarbeitung sensorischer Eingangssignale (Input), sensorische Integration, Verarbeitung motorischer Ausgangssignale (Output)
, sensorische Integration, Verarbeitung motorischer Ausgangssignale (Output)")
9
Sensorische Neurone übermitteln Information von den Augen und anderen Sinnesorganen, die externe Reize oder interne Zustände wahrnehmen Diese Information wird an Verarbeitungszentren im Gehirn oder in Ganglien gesandt, wo Interneurone den sensorischen Input integrieren Der motorische Output verlässt die Verarbeitungszentren über Motoneurone, die eine Reaktion hervorrufen, indem sie Muskeln oder Drüsen aktivieren

10
Viele Tierarten haben ein komplexes Nervensystem, das sich zusammensetzt aus:
einem Zentralnervensystem (ZNS), das die Integration bewerkstelligt; dazu gehören das Gehirn und ein Längsnervenstrang einem peripheren Nervensystem (PNS), das die Information zum ZNS hin- beziehungsweise von ihm wegleitet
, das die Integration bewerkstelligt; dazu gehören das Gehirn und ein Längsnervenstrang. einem peripheren Nervensystem (PNS), das die Information zum ZNS hin- beziehungsweise von ihm wegleitet.")
12
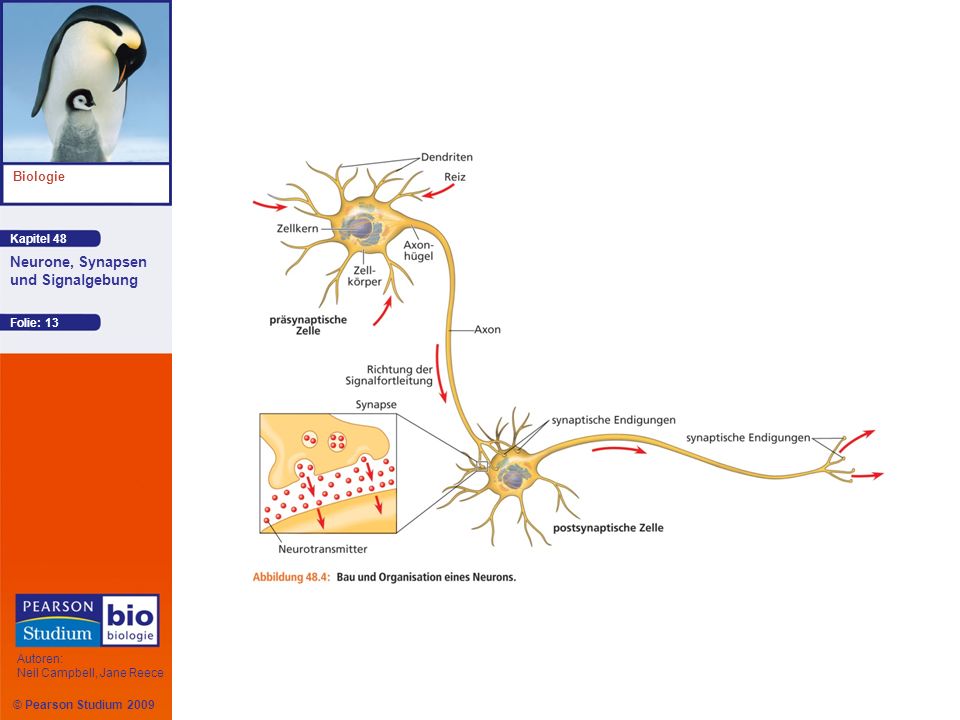
Neuronale Struktur und Funktion
Die meisten Organellen des Neurons liegen im Zellkörper Ein typisches Neuron weist zahlreiche Dendriten auf, stark verzweigte und sich dabei stetig verjüngende Fortsätze, die Signale von anderen Neuronen empfangen Ein Axon ist in der Regel viel länger als die Dendriten und leitet Signale an andere Zellen weiter Die kegelförmige Region eines Axons, wo es aus dem Zellkörper entspringt, wird als Axonhügel bezeichnet

14
Eine Synapse ist die Kontaktstelle zwischen einem Axon und einer anderen Zelle
Die synaptische Endigung eines Axons leitet über chemische Botenstoffe, so genannte Neurotransmitter, Information vom Senderneuron an das Empfängerneuron weiter

15
Das Senderneuron wird in Bezug auf die Synapse als präsynaptische Zelle, das Empfängerneuron als postsynaptische Zelle bezeichnet Die meisten Neuronen werden von Gliazellen ernährt

17
48.2: Aufrechterhaltung des Ruhepotenzials eines Neurons durch Ionenpumpen und Ionenkanäle
Alle Zellen haben ein Membranpotenzial, eine Spannung (elektrische Ladungsdifferenz) über ihrer Plasmamembran Veränderungen dieses Membranpotenzials wirken als Signale und leiten Information weiter Das Membranpotenzial eines ruhenden, nicht erregten Neurons ist das Ruhepotenzial
 über ihrer Plasmamembran. Veränderungen dieses Membranpotenzials wirken als Signale und leiten Information weiter. Das Membranpotenzial eines ruhenden, nicht erregten Neurons ist das Ruhepotenzial.")
18
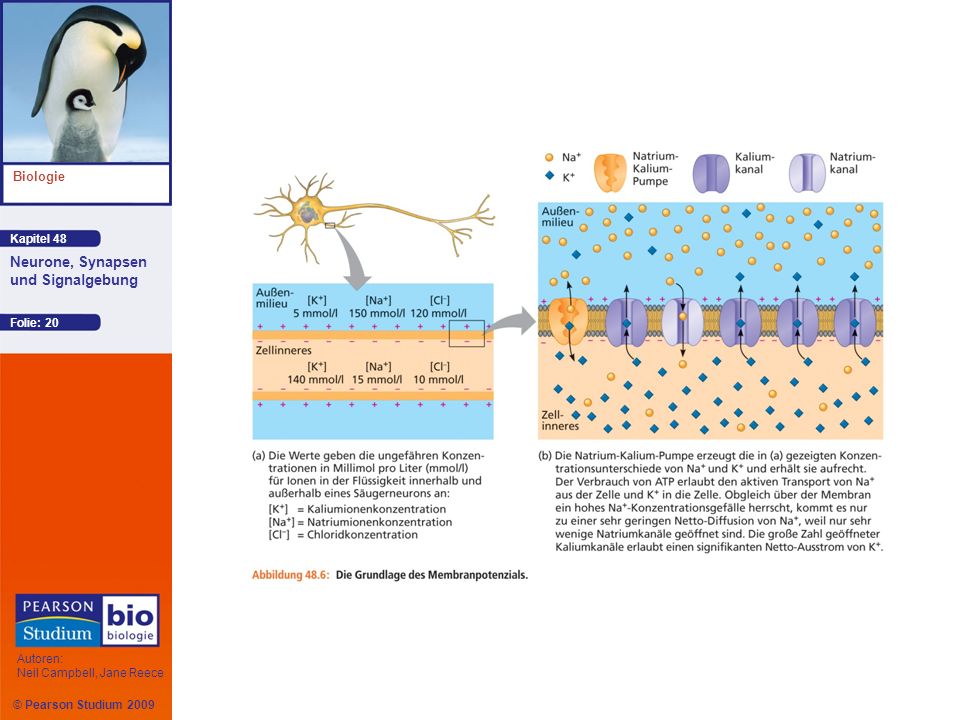
Entstehung des Ruhepotenzials
Beim Säugerneuron ist die K+-Konzentration im Inneren der Zelle höher, während die Na+-Konzentration im Außenmilieu höher ist Natrium-Kalium-Pumpen nutzen die Energie, die bei der Hydrolyse von ATP frei wird, um Na+ aktiv aus der Zelle und K+ in die Zelle zu transportieren Die Konzentrationsgradienten von K+ und Na+ über der Plasmamembran stellen eine chemische Form potenzieller Energie dar

19
Animation: Resting Potential
Ionenkanäle in der Plasmamembran wandeln das chemische Potenzial in ein elektrisches Potenzial um Ein Neuron in Ruhe weist weit mehr offene Kaliumkanäle auf als offene Natriumkanäle; K+ diffundiert durch offene Kaliumkanäle Intrazelluläre Anionen, die nicht durch die Membran ausströmen, tragen zu einem Überschuss von negativen Ladungen im Zellinneren bei Animation: Resting Potential

21
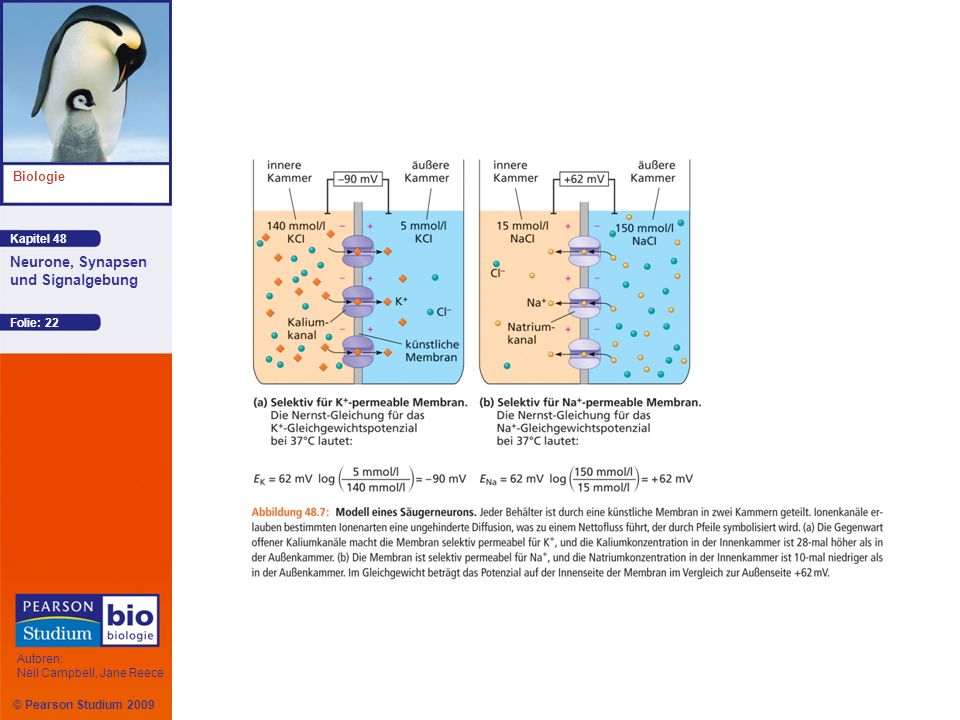
Ein Modell des Ruhepotenzials
Das Ruhepotenzial kann als simples Modell dargestellt werden, das aus zwei Kammern besteht, die durch eine künstliche Membran getrennt sind die Konzentration von Kaliumchlorid ist in der Innenkammer höher und geringer in der Außenkammer Kaliumionen wandern entsprechend ihrem Konzentrationsgradienten in die Außenkammer in der Innenkammer kommt es zu einem Überschuss an negativen Ladungen Wenn das Modellneuron sein Ionengleichgewicht erreicht hat, kompensiert der elektrische Gradient den chemischen Gradienten

23
Eion = 62 mV (log[ion]außen/[ion]innen)
Das Gleichgewichtspotenzial (Eion) ist die Größe der Membranspannung beim Gleichgewicht für ein bestimmtes Ion und lässt sich mithilfe der Nernst-Gleichung berechnen: Eion = 62 mV (log[ion]außen/[ion]innen) Das Gleichgewichtspotenzial von K+ (EK) ist negativ, wohingegen das Gleichgewichtspotenzial von Na+ (ENa) positiv ist
![Eion = 62 mV (log[ion]außen/[ion]innen)](http://slideplayer.org/slide/658014/1/images/23/Eion+%3D+62+mV+%28log%5Bion%5Dau%C3%9Fen%2F%5Bion%5Dinnen%29.jpg "Das Gleichgewichtspotenzial (Eion) ist die Größe der Membranspannung beim Gleichgewicht für ein bestimmtes Ion und lässt sich mithilfe der Nernst-Gleichung berechnen: Eion = 62 mV (log[ion]außen/[ion]innen) Das Gleichgewichtspotenzial von K+ (EK) ist negativ, wohingegen das Gleichgewichtspotenzial von Na+ (ENa) positiv ist.")
24
In einem ruhenden Neuron entspricht der Konzentrationsgradient für Na+ demjenigen für K+ und die Ionenkonzentrationen auf beiden Seiten der Membran bleiben weitgehend konstant

25
48.3: Axonale Fortleitung von Aktionspotenzialen
Neurone besitzen gesteuerte Ionenkanäle, die sich in Folge eines Reizes öffnen oder schließen können

27
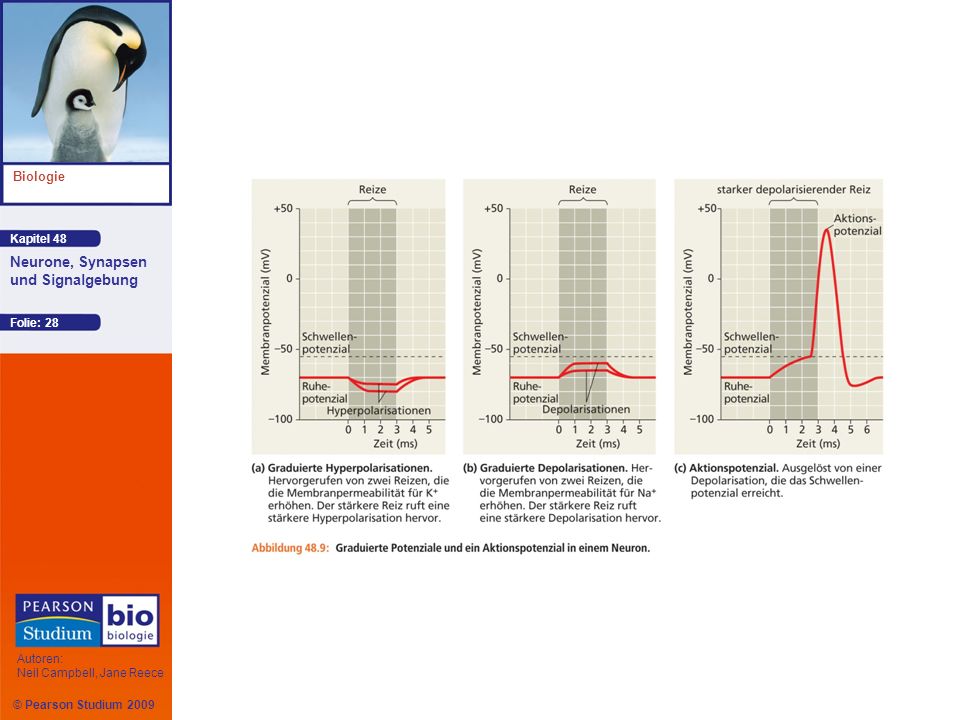
Das Öffnen oder Schließen von Ionenkanälen verändert das Membranpotenzial
Das Öffnen zusätzlicher Kaliumkanäle erhöht die Nettodiffusion von K+ aus dem Neuron heraus und die Innenseite der Membran wird negativer Diesen Vorgang, bei dem sich das Membranpotenzial EK (–90m bei 37°C) nähert, bezeichnet man als Hyperpolarisation
 nähert, bezeichnet man als Hyperpolarisation.")
29
Die Verringerung der Größe des Membranpotenzials wird als Depolarisation bezeichnet
An der Depolarisation von Neuronen sind häufig gesteuerte Natriumkanäle beteiligt

31
Aktionspotenziale treten immer dann auf, wenn eine Depolarisation die Membranspannung auf einen bestimmten Wert, das Schwellenpotenzial, anhebt Da Aktionspotenziale entweder in voller Größe auftreten oder gar nicht, spricht man von einer Alles-oder-Nichts-Reaktion auf Reize Aktionspotenziale sind die Nervenimpulse oder -signale, die Information über ein Axon weiterleiten

32
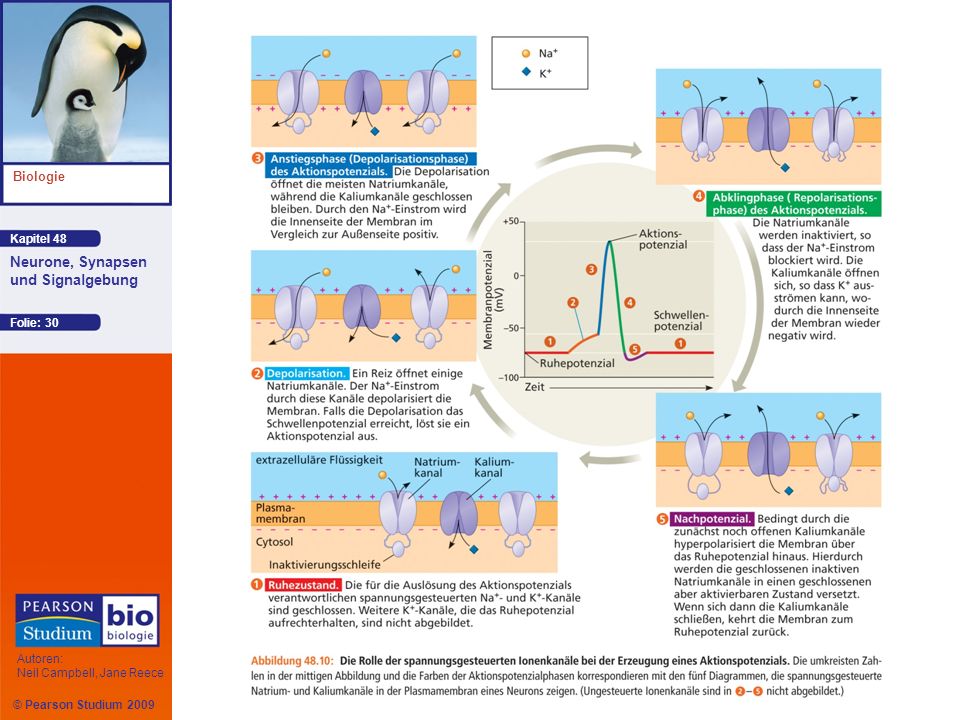
Erzeugung von Aktionspotenzialen: Eine nähere Betrachtung
Ein Neuron kann Hunderte von Aktionspotenzialen pro Sekunde erzeugen Die Frequenz, mit der ein Neuron Aktionspotenziale generiert, übermittelt Information über die Signalstärke Ein Aktionspotenzial kann in eine Folge von Phasen zerlegt werden

33
Beim Ruhepotenzial sind die meisten spannungsgesteuerten Natriumkanäle geschlossen, aber einige Kaliumkanäle sind offen

34
Wenn ein Reiz die Membran depolarisiert, öffnen sich einige spannungsgesteuerte Natriumkanäle, so dass mehr Na+ in die Zelle einströmen kann Während der Anstiegsphase wird das Schwellenpotenzial überschritten, was die Membran rasch in die Nähe von ENa bringt Während der Abklingphase werden die spannungsgesteuerten Natriumkanäle inaktiviert, während sich die meisten spannungsgesteuerten Kaliumkanäle öffnen, was einen raschen Kaliumausstrom zur Folge hat

35
Beim Nachpotenzial (undershoot) ist die Membranpermeabilität für K+ kurzfristig höher als in Ruhe, daher liegt das Membranpotenzial in dieser Phase näher an EK als das Ruhepotenzial; der Inaktivierungszustand wird überwunden
 ist die Membranpermeabilität für K+ kurzfristig höher als in Ruhe, daher liegt das Membranpotenzial in dieser Phase näher an EK als das Ruhepotenzial; der Inaktivierungszustand wird überwunden")
36
BioFlix: How Neurons Work Animation: Action Potential
Während der Refraktärzeit oder Refraktärperiode nach einem Aktionspotenzial kann kein zweites Aktionspotenzial ausgelöst werden Die Refraktärzeit ist eine Folge der Inaktivierung von Natriumkanälen BioFlix: How Neurons Work Animation: Action Potential

37
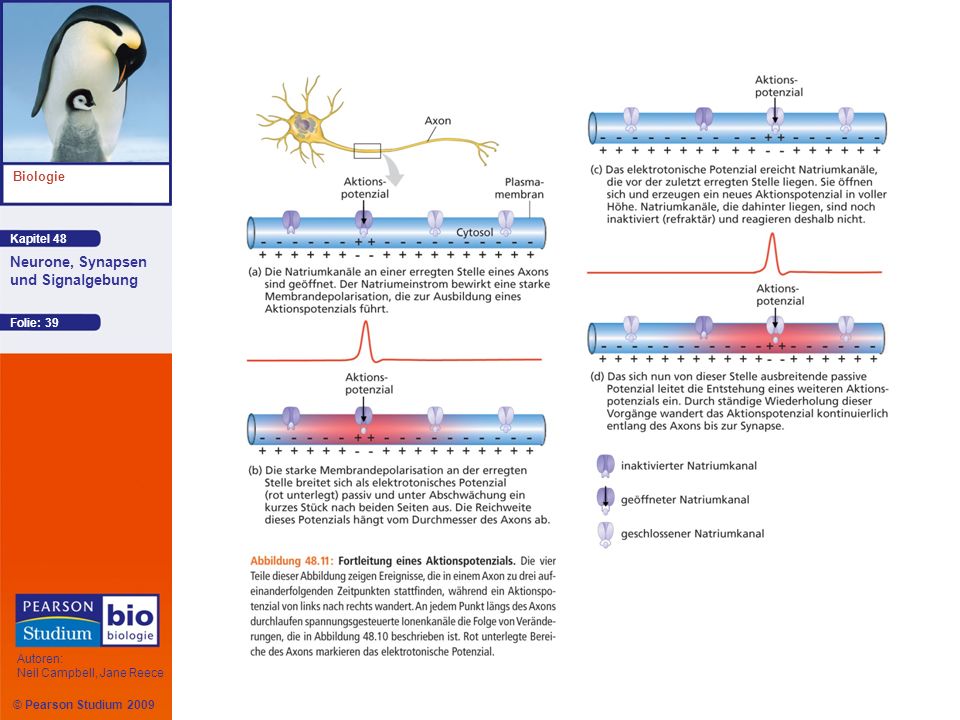
Fortleitung von Aktionspotenzialen
Ein Aktionspotenzial pflanzt sich über weite Strecken fort, weil es sich lokal immer wieder selbst regeneriert An dem Ort, an dem ein Aktionspotenzial ausgelöst wird, generiert der Natriumeinstrom während der Depolarisationsphase einen elektrischen Strom, der die benachbarten Regionen der Axonmembran depolarisiert

38
Da unmittelbar hinter jedem lokalen Aktionspotenzial eine Zone liegt, in der die Natriumkanäle auf Grund des dort zuvor erzeugten Aktionspotenzials noch inaktiviert sind, wird verhindert, dass Aktionspotenziale zurück zum Zellkörper wandern Aktionspotenziale bewegen sich nur in eine Richtung – hin zu den synaptischen Endigungen

40
Fortleitungsgeschwindigkeit
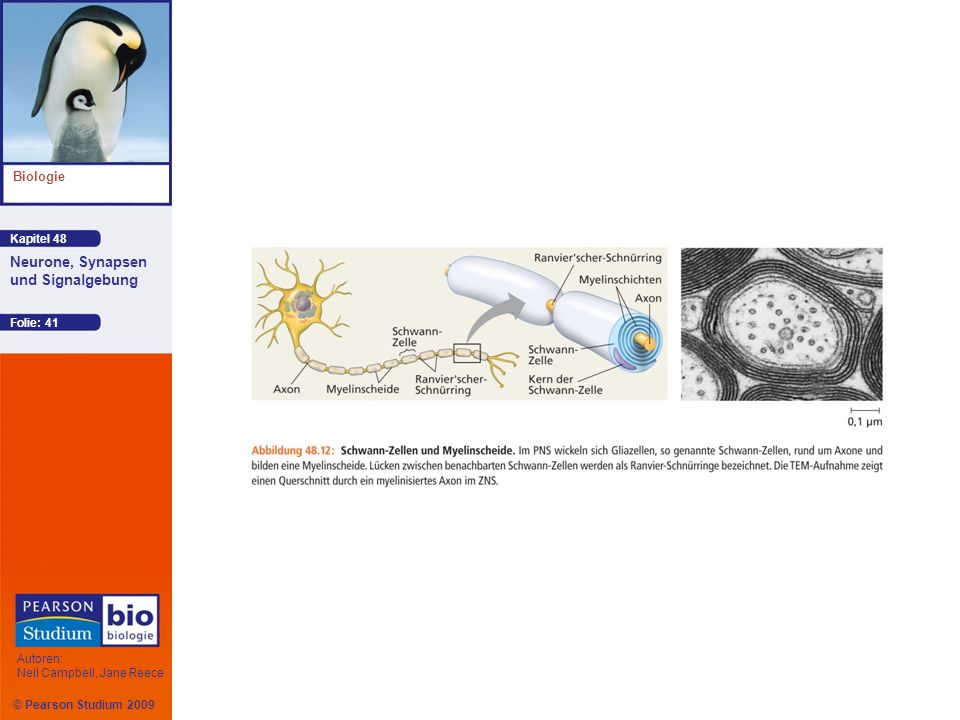
Die Geschwindigkeit eines Aktionspotenzials steigt mit dem Durchmesser des Axons Die Axone von Vertebraten sind von einer Myelinscheide umhüllt, was die rasche Fortleitung in dünnen Axonen ermöglicht Die Myelinscheiden oder -hüllen werden von zwei Gliazelltypen erzeugt – von Oligodendrocyten im ZNS und von Schwann-Zellen im PNS

42
Bei einem myelinisierten Axon beschränken sich spannungsgesteuerte Natriumkanäle auf Lücken in der Myelinscheide, die als Ranvier-Schnürringe bezeichnet werden Das Aktionspotenzial “springt” von Schnürring zu Schnürring, ein Mechanismus, der als saltatorische Erregungsleitung bezeichnet wird

44
48.4: Synapsen als Kontaktstellen zwischen Neuronen
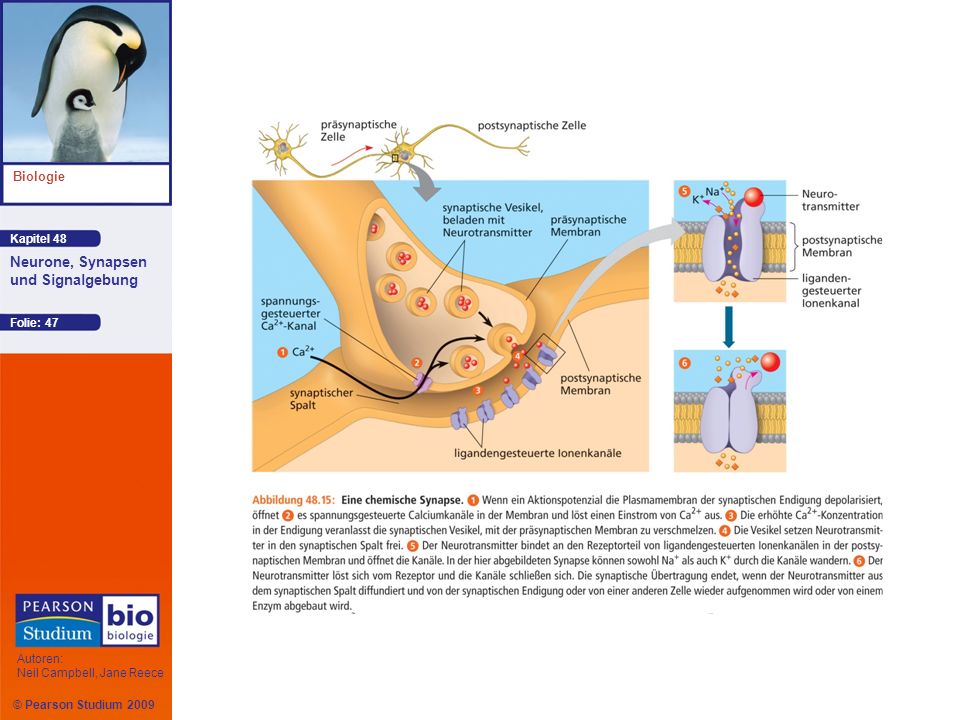
Bei elektrischen Synapsen kann der elektrische Strom schnell und in beiden Richtungen von einem Neuron zum anderen fließen Bei chemischen Synapsen setzt das präsynaptische Neuron chemische Neurotransmitter frei Die meisten Synapsen sind chemische Synapsen

46
An jeder präsynaptischen Endigung liegt der jeweilige Neurotransmitter in zahlreichen Vesikeln verpackt vor, die als synaptische Vesikel bezeichnet werden Das Eintreffen eines Aktionspotenzials verursacht die Freisetzung der Neurotransmitter Die Neurotransmittermoleküle diffundieren dann durch den synaptischen Spalt zum postsynaptischen Neuron Animation: Synapse

48
Erzeugung postsynaptischer Potenziale
Bindet ein Neurotransmitter an den extrazellulären Teil eines ligandengesteuerten Ionenkanals, so öffnet sich der Kanal und erlaubt bestimmten Ionen, durch die postsynaptische Membran zu diffundieren Bindet ein Neurotransmitter an einen Kanaltyp, der sowohl K+ als auch Na+ passieren lässt, öffnen sich die Kanäle und ein postsynaptisches Potenzial entsteht

49
Postsynaptische Potenziale lassen sich in zwei Katogerien unterteilen:
erregende (exzitatorische) postsynaptische Potenziale (EPSPs) sind Depolarisationen, die das Membranpotenzial in Richtung Schwellenpotenzial verschieben hemmende (inhibitorische) postsynaptische Potenziale (IPSPs) sind Depolarisationen, die das Membranpotenzial vom Schwellenpotenzial wegschieben
 postsynaptische Potenziale (EPSPs) sind Depolarisationen, die das Membranpotenzial in Richtung Schwellenpotenzial verschieben. hemmende (inhibitorische) postsynaptische Potenziale (IPSPs) sind Depolarisationen, die das Membranpotenzial vom Schwellenpotenzial wegschieben.")
50
Nach ihrer Freisetzung können Neurotransmitter
sich durch einfache Diffusion aus dem synaptischen Spalt entfernen in synaptische Vesikel verpackt oder in Gliazellen transportiert werden von einem Enzym abgebaut werden

51
Summation postsynaptischer Potenziale
Im Gegensatz zu Aktionspotenzialen sind postsynaptische Signale abgestuft und regenerieren sich nicht Die meisten Synapsen auf einem Neuron sitzen auf seinen Dendriten oder seinem Zellkörper Ein einzelnes EPSP ist gewöhnlich zu klein, um in einem postsynaptischen Neuron ein Aktionspotenzial auszulösen Wenn zwei EPSPs rasch hintereinander an derselben Synapse eintreffen, tritt ein Vorgang ein, den man als zeitliche Summation bezeichnet

53
Bei der zeitlichen Summation, summieren sich EPSPs, die fast gleichzeitig von verschiedenen Synapsen generiert werden Die Kombination vieler EPSPs mittels zeitlicher und räumlicher Summation kann ein Aktionspotenzial generieren

54
Durch Summation kann ein IPSP einem EPSP entgegenwirken
Die addierten Effekte sämtlicher EPSPs und IPSPs bestimmen, ob am Axonhügel das Schwellenpotenzial erreicht und ein Aktionspotenzial ausgelöst wird

55
Modulation der synaptischen Übertragung
Bei Second-Messenger-Systemen bindet ein Neurotransmitter an einen Rezeptor, der kein Teil des Ionenkanals ist Diese Bindung aktiviert einen Signalübertragungsweg, an dem ein sekundärer Botenstoff beteiligt ist Die Wirkung der Second-Messenger-Systeme setzt langsamer ein, hält dafür aber länger an

56
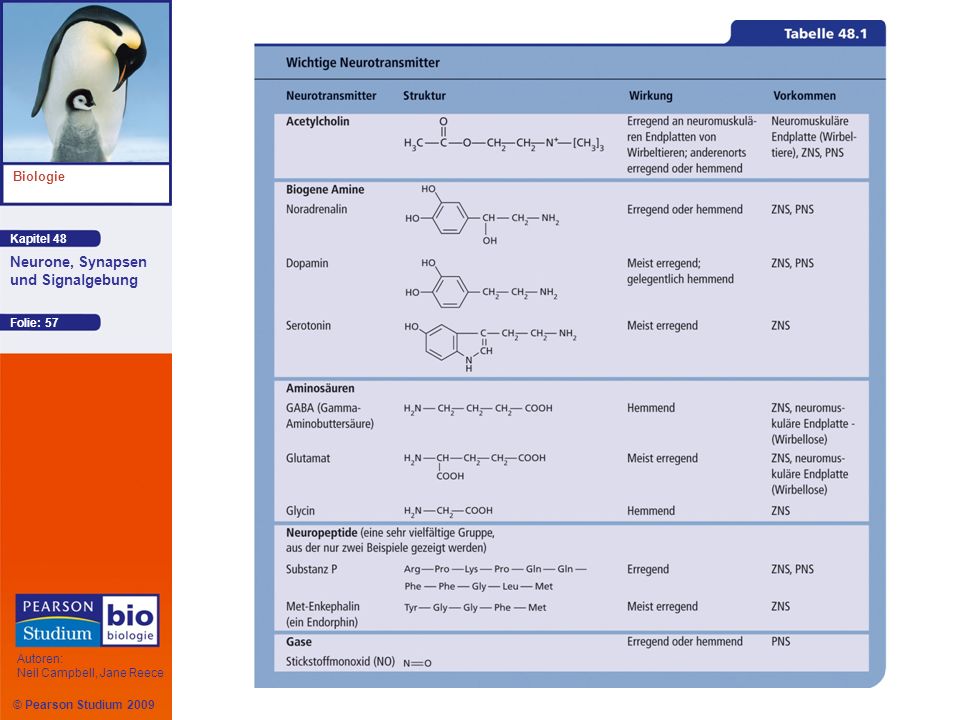
Neurotransmitter Ein einzelner Neurotransmitter kann sich wesentlich in seiner Wirkung auf postsynaptische Zellen unterscheiden Die Hauptklassen, in die Neurotransmitter fallen, sind Acetylcholin, biogene Amine, Aminosäuren, Neuropeptide und Gase

58
Acetylcholin Acetylcholin ist einer der häufigsten Neurotransmitter bei Vertebraten und Evertebraten Bei Vertebraten ist es für gewöhnlich ein erregender Transmitter

59
Biogene Amines Zu den biogenen Aminen gehören Adrenalin, Noradrenalin, Dopamin und Serotonin Sie sind im ZNS und im PNS aktiv

60
Aminosäuren Im ZNS von Vertebraten spielen zwei Neurotransmitter eine wichtige Rolle: Gamma-Aminobuttersäure (GABA) und Glutamat
 und Glutamat.")
61
Neuropeptide Mehrere Neuropeptide, aus wenigen Aminosäuren bestehende Oligopeptide, wirken ebenfalls als Neurotransmitter Zu den Neuropeptiden gehören Substanz P und die Endorphine, die beide auf das Schmerzempfinden wirken Da Opiate an dieselben Rezeptorproteine wie Endorphine binden, rufen sie dieselben physiologischen Effekte hervor

Ähnliche Präsentationen















 U N I V E R S I T Ä T H A M B U R G November 2011.>")
Media Landesanstalt für Kommunikation Baden-Württemberg (LFK) Landeszentrale für Medien und Kommunikation.>")











