Präsentation herunterladen
Die Präsentation wird geladen. Bitte warten

1
Kap. 4.1 Datenstruktur und Eigenschaften
Kap. 4 Positionsbäume Kap Datenstruktur und Eigenschaften Zweck:z.B. Textverarbeitung., genetische Information (Genom 109B) x = x1x2 ... xn X*; xi X (X ist Alphabet) Leerzeichen; leere Zeichenreihe n sehr groß, z.B. 107, wie groß für großen Brockhaus? y = y1, ..., ym Def.: y kommt in x an Stelle i vor, wenn xi xi xi+m-1 = y
 x = x1x2 ... xn X*; xi X (X ist Alphabet) Leerzeichen; leere Zeichenreihe. n sehr groß, z.B. 107, wie groß für großen Brockhaus y = y1, ..., ym. Def.: y kommt in x an Stelle i vor, wenn xi xi xi+m-1 = y.")
2
(P1) Finde alle Vorkommen von y in x,
z.B. für Buchindex ersetze überall INHALT durch VOLUMEN 1. Verwende y, um Textstellen zu identifizieren 2. Wird Stelle durch u eindeutig identifiziert? „Positionsidentifikator“ Def.: u heißt Positionsidentifikator für Pos. i in x, wenn u kürzeste, eindeutige Teilzeichenreihe von x beginnend in Pos. i ist, d.h. x = yuz x = y`uz ` y = y` und u ist kürzeste Teilzeichenreihe mit dieser Eigenschaft. Hinw.: |y| = i-1, z = z` Problem: u existiert nicht immer, z.B. bbb, Ident. für Pos.2?

3
Lemma: Sei $ Sonderzeichen, d.h. $ X,
dann hat jede Position in x$ einen Positions-Identifikator (Umkehrung?) Bew.: In x1x2 ... xu$ kommt xi ... xu$ genau einmal vor, jetzt kürze xi ... xu$ sukzessive von hinten Beispiel: abab $ p(1) = aba; p(2) = ba; p(3) = ab$ p(4) = b $; p(5) = $ Def.: X-Baum: orientierter oder geordneter Baum mit Kantenmarken (Selektoren) aus X
 Bew.: In x1x2 ... xu$ kommt. xi ... xu$ genau einmal vor, jetzt kürze xi ... xu$ sukzessive von hinten. Beispiel: abab $ p(1) = aba; p(2) = ba; p(3) = ab$ p(4) = b $; p(5) = $ Def.: X-Baum: orientierter oder geordneter Baum mit Kantenmarken. (Selektoren) aus X.")
4
Def.: Positionsbaum T für
x$ = x1x2 ... xu $ mit xi X ist ein X {$}-Baum mit: 1. T hat u + 1 Blätter markiert mit 1, 2, ..., u + 1 für Positionen von x$ 2. Markenfolge längs Pfad von Wurzel zu Blatt i ist Pos.Id. p(i) für i Problem: Invarianz von Pos.Ids., z.B. (Kap., Unterkap., Absatz, Wort) Lemma: x$ hat eindeutigen Positions-Baum Bew.: Eindeutigkeit der Pos.Ids. in x$
 für i. Problem: Invarianz von Pos.Ids., z.B. (Kap., Unterkap., Absatz, Wort) Lemma: x$ hat eindeutigen Positions-Baum. Bew.: Eindeutigkeit der Pos.Ids. in x$")
5
Beispiel: Für abab$ 1 2 3 4 5 a b $

6
Kap. 4.2 Konstruktions-Algorithmen
Lemma: Falls x , dann hat jedes Blatt von x $ mindestens einen Bruder Bew.: d.h. p(i) = ... ab ist kürzeste identifizierende Teilzeichenreihe für Pos. i. Wenn keinen Bruder hätte, könnte man p(i) kürzen, siehe auch Beispiel für abab$ i b a i
 = ... ab. ist kürzeste identifizierende Teilzeichenreihe. für Pos. i. Wenn keinen Bruder hätte, könnte. man p(i) kürzen, siehe auch Beispiel für abab$ i. b. a. i.")
7
4.2.1 Naive Konstruktion D A T E N S T R U K T U R E N $
1 2 3 4 . Hinw.: i.e. Algorithmus (n2) Bibel 100 MB = 108 Bytes Beobachtung: kurze Pos. Identifikatoren
 Bibel 100 MB = 108 Bytes. Beobachtung: kurze Pos. Identifikatoren.")
8
4.2.2 Alg. für Rechts-links Konstruktion
von Pos. Baum Ti (xi xi xn$) konstruiere Ti-1 (xi-1 xi ... xn$) d.h. in Ti folge Pfad mit Marken: xi-1 xi ... bis Fall a: Blatt j erreicht wird, d.h. p(j) kommt auch an Stelle i-1 vor p(j), p(i-1) verlängern j
 konstruiere Ti-1 (xi-1 xi ... xn$) d.h. in Ti folge Pfad mit Marken: xi-1 xi ... bis. Fall a: Blatt j erreicht wird, d.h. p(j) kommt auch an Stelle i-1 vor. p(j), p(i-1) verlängern. j.")
9
Fall b: Zwischenknoten z erreicht wird, von dem aus p(i-1) nicht
weiterführt z . . . Beispiel: T (abba$) 12345 T5: T4: $
 T5: T4: $")
10
Ordnung nach Kantenmarken für schnellen Selektor-Zugriff!
Aufwand: mit Kostenamortisierung Nachteil: Konstruktion von T(x $) nach voller Eingabe von x. T für umgekehrten Text? x = x1 x2 ... xn x$ x = xn xn x x$ xi ... x2x1$ p`(i) Textstellen zur Identifizierung rückwärts eingeben
 nach voller Eingabe von x. T für umgekehrten Text x = x1 x2 ... xn x$ x = xn xn x1 x$ xi ... x2x1$ p`(i) Textstellen zur Identifizierung rückwärts eingeben.")
11
„on-line“ Verfahren, Konstruktion von T(x) in Realzeit mitlaufend.
Links-Rechts Konstr. „on-line“ Verfahren, Konstruktion von T(x) in Realzeit mitlaufend. Triviale Idee: Übergang von T(x1 ... xi$) T (x1 ... xi xi+1$) Änderungen in T an vielen Stellen Def.: Partieller Pos. Baum PJ(x1...xi) mit J [1:i] enthält für k J genau kürzeste Zeichenreihe ab k: pJ(k), die Position k innerhalb von J identifiziert, (d.h. pJ(k) kommt an keiner anderen Stelle m J vor. Anm.: PJ(x) existiert nicht immer
 in Realzeit mitlaufend. Triviale Idee: Übergang von. T(x1 ... xi$) T (x1 ... xi xi+1$) Änderungen in T an vielen Stellen. Def.: Partieller Pos. Baum PJ(x1...xi) mit J [1:i] enthält für k J. genau kürzeste Zeichenreihe ab k: pJ(k), die Position k innerhalb von J identifiziert, (d.h. pJ(k) kommt an keiner anderen Stelle m J vor. Anm.: PJ(x) existiert nicht immer.")
12
Übergang: P[1:3](x) zu P[1:4](x)
Beispiel: x = abbab P{1}(x) = P{1,2}(x) = a b 1 2 P{2,3}(x) = P[1:3](x) = P{1,4}(x) 3 Übergang: P[1:3](x) zu P[1:4](x) x = abbab P[1:4](x) existiert nicht P[1:4](xa) 1 2 3 a b P[1 : 5](abbaba) P[1 : 5](abbaba$) Jede Position hat Identifikator
 zu P[1:4](x)](http://slideplayer.org/slide/207036/1/images/12/%C3%9Cbergang%3A+P%5B1%3A3%5D%28x%29+zu+P%5B1%3A4%5D%28x%29.jpg "Beispiel: x = abbab. P{1}(x) = P{1,2}(x) = a. b P{2,3}(x) = P[1:3](x) = P{1,4}(x) 3. Übergang: P[1:3](x) zu P[1:4](x) x = abbab. P[1:4](x) existiert nicht. P[1:4](xa) a. b. P[1 : 5](abbaba) P[1 : 5](abbaba$) Jede Position hat Identifikator.")
13
Fortschreibung von P[1:4] (abbaba) zu P[1:5] (abbaba$)
2 3 1 4 Lemma: Für n = |x| gilt: P[1: n+1](x$) = T(x$) Bew.: nach Konstruktion, es werden genau die Pos. Identifikatoren eingefügt
![Fortschreibung von P[1:4] (abbaba) zu P[1:5] (abbaba$)](http://slideplayer.org/slide/207036/1/images/13/Fortschreibung+von+P%5B1%3A4%5D+%28abbaba%29+zu+P%5B1%3A5%5D+%28abbaba%24%29.jpg "Lemma: Für n = |x| gilt: P[1: n+1](x$) = T(x$) Bew.: nach Konstruktion, es werden genau die Pos. Identifikatoren. eingefügt.")
14
Monotonie Lemma für Pos. Identifikatioren:
Sei e(i) die Pos., bei der p(i) endet, dann gilt: i < j e(i) e(j) Bew.: Trivial, falls i = e(i), sonst Bew. durch Widerspruch: Sei i < j und e(i) > e(j) p(i) x i j e(j) e(i) p(j) p(i) = xi xi xe(i) d.h. xi ... xe(i) kommt nochmals vor d.h. p(j) kommt auch nochmals vor, ist nicht Pos.Id.
 die Pos., bei der p(i) endet, dann gilt: i < j e(i) e(j) Bew.: Trivial, falls i = e(i), sonst Bew. durch Widerspruch: Sei i < j und e(i) > e(j) p(i) x. i. j. e(j) e(i) p(j) p(i) = xi xi xe(i) d.h. xi ... xe(i)-1 kommt nochmals vor. d.h. p(j) kommt auch nochmals vor, ist nicht Pos.Id.")
15
Graphische Deutung: ... xi xi+1 ... xk ... deren Pos.Ids.
hier enden Intervall von Positionen z.B. a b a $ a b b

16
d.h. bei links-rechts Aufbau von T(x)
kann für i < j p(j) frühestens mit p(i) entstehen. Grundidee: für links – rechts Konstruktion des Positions-Baums: Für x1x2 ... xi konstruiere partiellen Pos.B. für alle Positionen mit Pos.Ids. einschl der in i endenden x1x xk xk ... xi d.h. P[1:k-1]
 frühestens mit p(i) entstehen. Grundidee: für links – rechts Konstruktion des Positions-Baums: Für x1x2 ... xi konstruiere partiellen Pos.B. für alle. Positionen mit Pos.Ids. einschl der in i endenden. x1x2 ... xk-1 xk ... xi. d.h. P[1:k-1]")
17
xk ... xi ist Anfang von p(k),
entspricht Pfad durch P[1:k-1] (x1 ... xi) Führe Marke in P[1:k-1] (x1 ... xi) k xk xi k Fortschreibung von P[1:k-1] (x1 ... xi) bei Eingabe von xi+1:
 Führe Marke in P[1:k-1] (x1 ... xi) k. xk. xi. k. Fortschreibung von P[1:k-1] (x1 ... xi) bei Eingabe von xi+1:")
18
Fall 1: neben internem Knoten
1.1 verschiebe längs vorhandener Kante xi+1 weiter nach unten wird über neue Kante xi+1 zu Blatt , p(k) fertig. Führe ein, iteriere Fall 2: steht neben Blatt l: verlängere p(l) um 1 Zeichen y, neues Blatt k k k k k+1 k l l xi+1 y = xi+1 : l l iteriere für 2.2 y xi+1 : xi+1 y k+1 l k
 fertig. Führe ein, iteriere. Fall 2: steht neben Blatt l: verlängere p(l) um 1 Zeichen y, neues Blatt. k. k. k. k. k+1. k. l. l. xi y = xi+1 : l. l. iteriere für. 2.2 y xi+1 : xi+1. y. k+1. l. k.")
19
Eintrag von Blatt , schrittweiser Aufbau von p(k), gesamt |p(k)|
1.1 1.2 k k 2.1 2.2 k l k l Kosten: Eintrag von Blatt , schrittweiser Aufbau von p(k), gesamt |p(k)| für ganz T(x $): k | x | + 1 c | p ( i ) | å i = 1
, gesamt |p(k)| für ganz T(x $): k. | x. | + 1. c. | p. ( i. ) | å. i. = 1.")
20
Für sprachliche Texte typisch:
|p(i)| ~ log |x| Gesamtkosten (|x|log|x|) wie bei rechts - links Konstruktion! Praktische Variante: partielle Pos.B. für Positionen entsprechend Wortanfängen, nach , CR, Seitenwechsel, etc. oder entsprechend Teilsatzanfängen (z.B. Lexikon für Zitate) nach. ; ! ? : etc.
| ~ log |x| Gesamtkosten (|x|log|x|) wie bei rechts - links Konstruktion! Praktische Variante: partielle Pos.B. für Positionen entsprechend Wortanfängen, nach , CR, Seitenwechsel, etc. oder entsprechend Teilsatzanfängen (z.B. Lexikon für Zitate) nach. ; ! : etc.")
21
4.2.4 Korrektur von Texten und zugehörigen Positions-Bäumen
Hauptproblem: Invarianz der Positionen, verwende Pos. #, die gegenüber typischen Änderungen weitgehend invariant sind, 4.20 für Folie 20 in Kap. 4 Korrekturoperationen: streichen: x = u v w z u, z : invariant; v : streichen; w : variant Ansatz: lösche vw; füge w mit neuen Pos. ein

22
2. einfügen: x = u w z v einfügen zu x` = u v w z u v w z wie vorher u w z wie vorher Ansatz: lösche w; füge v w mit neuer Position ein 3. ersetzen: v durch v`in x = u v w z zu x`= u v`w z Ansatz: lösche v w; füge v`w mit neuen Positionen ein Grundidee für Alg.: Schritt 1: bestimme y = v w in 1. und 3. y = w in 2. Schritt 2: delete (y, x) in x = u y z durch Übergang von T(x) zu partiellem Pos.Baum ohne diejenigen Positionen j, deren p(j) mit y überlappen.
 in x = u y z. durch Übergang von T(x) zu partiellem Pos.Baum. ohne diejenigen Positionen j, deren p(j) mit y überlappen.")
23
Schritt 3: Sei y`, w, v w, v`w entsprechend 1., 2., 3.
Ergänze partiellen Pos.Baum aus Schritt 2 zu T(x`); x`= u y`z zu Schritt 2: delete y in u y z, welche p(j) überlappen y? u1u2 ... uk-1 yk ... ylz Menge der Positionen j, deren p(j) zu entfernen sind
; x`= u y`z. zu Schritt 2: delete y in u y z, welche p(j) überlappen y u1u2 ... uk-1 yk ... ylz. Menge der Positionen j, deren p(j) zu entfernen sind.")
24
Def.: A(j) = {i: e(i) = j}, Menge von Anfangspositionen def
Anm.: nach Monotonie-Lemma ist A(j) ein Intervall y k l x p(j) j e(j) 1. e(j) < k keine Überlappung 2. j > l keine Überlappung (1. 2.) = = e(j) k j l Überlappung ~ A(k,l) = {j : j l e(j) k} def
 ein Intervall. y. k. l. x. p(j) j. e(j) 1. e(j) < k keine Überlappung. 2. j > l keine Überlappung. 3. (1. 2.) = 1. 2. = e(j) k j l Überlappung. ~ A(k,l) = {j : j l e(j) k} def.")
25
Überlegung: A(k,l) ist Intervall von Positionen:
~ Überlegung: A(k,l) ist Intervall von Positionen: min(j) : l e(j) k Def.: T(x$)\ J = P[1:|x|+1]\J (x$) def Lemma: Sei x = u y z; u`= u y`z y k l y` m ~ T(x$)\A(k,l) = T(x`$)\A(k,m)
 ist Intervall von Positionen: min(j) : l. e(j) k. Def.: T(x$)\ J = P[1:|x|+1]\J (x$) def. Lemma: Sei x = u y z; u`= u y`z. y. k. l. y` m. ~ T(x$)\A(k,l) = T(x`$)\A(k,m)")
26
Bew.: Nach Konstruktion
Korrektur Alg.: ersetze y durch y` T(x$) sei vorhanden 1. Übergang von T(x$) T(x$)\A(k,l) 2. ersetze Text x = u y z x`= u y`z 3. Ergänze T(x`$)\A(k,m) zu T(x´$) ~ ~ Korrektur-Kosten: pro Entfernung von p(j) Aufwand |p(j)| log|x|, ebenso pro Einfügung (|A(k,l) + |A(k,m)|) •log|x|
 sei vorhanden. 1. Übergang von T(x$) T(x$)\A(k,l) 2. ersetze Text x = u y z x`= u y`z. 3. Ergänze T(x`$)\A(k,m) zu T(x´$) ~ ~ Korrektur-Kosten: pro Entfernung. von p(j) Aufwand |p(j)| log|x|, ebenso pro Einfügung. (|A(k,l) + |A(k,m)|) •log|x|")
27
4.3 Anwendungen von Pos. Bäumen
für Texte und genetische Information: (P1) Finde alle Vorkommen von y in x y
 Finde alle Vorkommen von y in x. y.")
28
(P3) Finde längste mehrfach vorkommende Teilzeichenreihe:
(P2) Text Korrektur (P3) Finde längste mehrfach vorkommende Teilzeichenreihe: Führe Pegel für tiefste Blätter bei Baumaufbau Beispiel: Haben Programme X, größere gemeinsame Teile? (P4) Interne Mustergleichheit: längste Zeichenreihe ab Pos. i, die mehrfach vorkommt? Länge = |p(i)|-1 an Positionen in diesem Unterbaum i (P5) Muster in DNA-Ketten
 Text Korrektur. (P3) Finde längste mehrfach vorkommende Teilzeichenreihe: Führe Pegel für tiefste Blätter bei Baumaufbau. Beispiel: Haben Programme X, größere gemeinsame Teile (P4) Interne Mustergleichheit: längste Zeichenreihe ab Pos. i, die mehrfach vorkommt Länge = |p(i)|-1 an Positionen. in diesem Unterbaum. i. (P5) Muster in DNA-Ketten.")
29
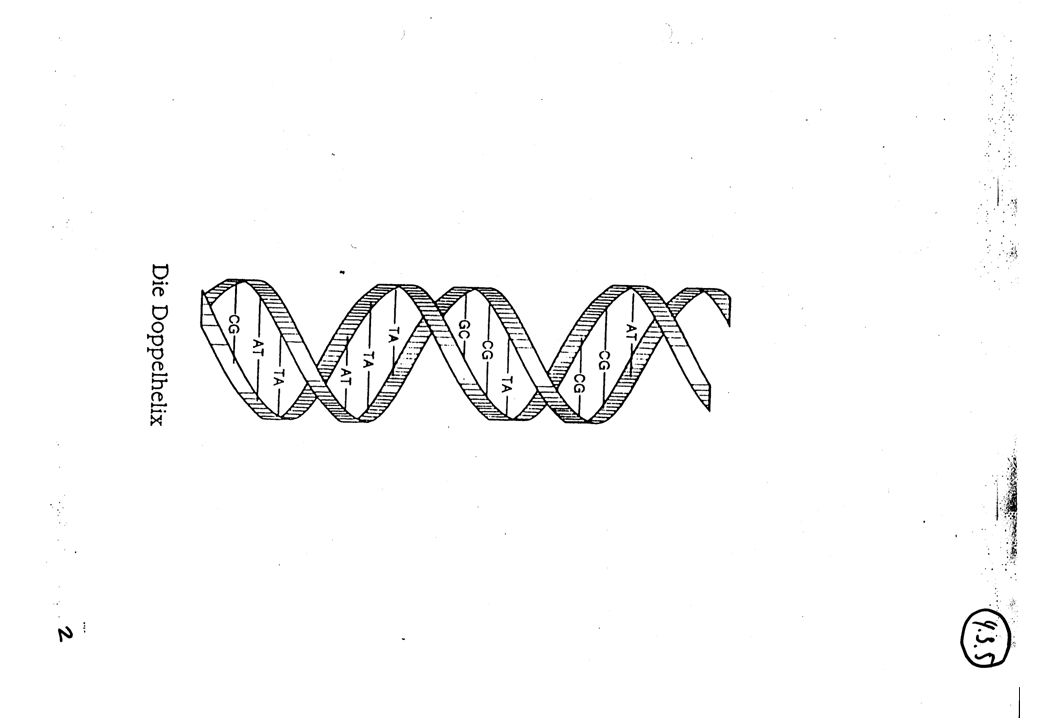
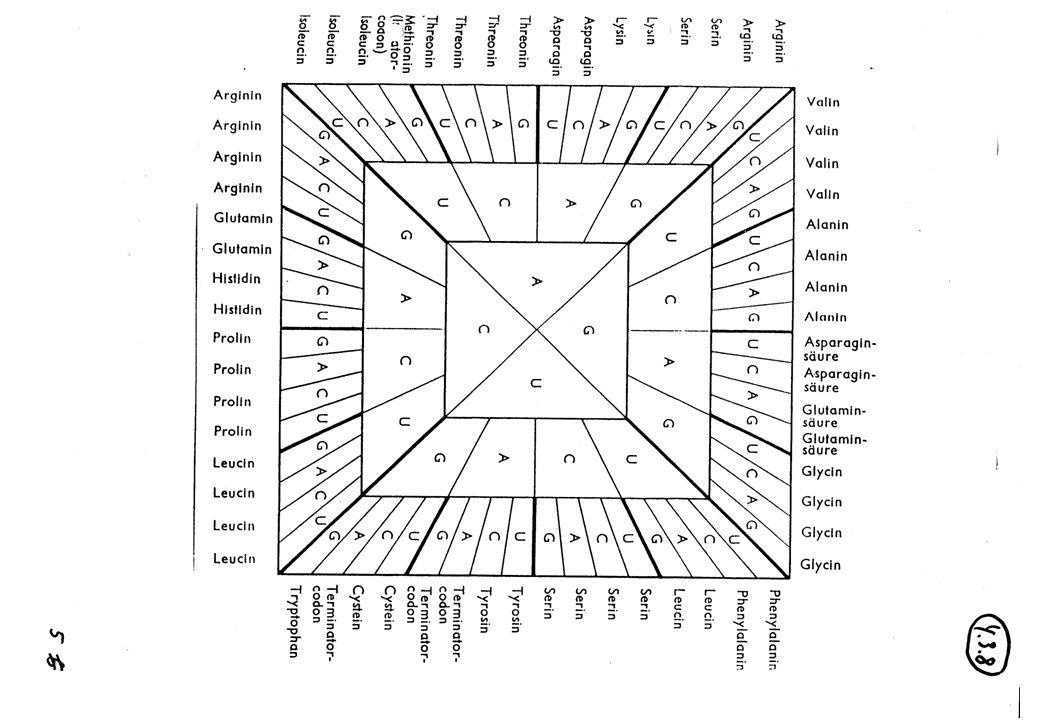
Anwendung Genom Datenbanken
U : Uracil C : Cytosin A : Adenin G : Guanin Basen von Nukleotid-Bausteinen in Messenger-RNS Triplett codiert den Einbau einer von 20 Aminosäuren bei Proteinsynthese, z.B. UGG ~ Tryptophan (= Aminosäure) Einbau 43 Kombinationen = 64 Zeichen im Grundalphabet von Tripletts RNS-Sequenzen von Trippletts ~ Aminosäuren-Sequenzen in Proteinmolekülen (Primärstruktur) Problem: Finde gesuchte RNS-Sequenzen innerhalb von bekannten RNS-Sequenzen typisch 104 bis 105 Tripletts pro Protein
 Einbau. 43 Kombinationen = 64 Zeichen im Grundalphabet von Tripletts. RNS-Sequenzen von Trippletts ~ Aminosäuren-Sequenzen in Proteinmolekülen (Primärstruktur) Problem: Finde gesuchte RNS-Sequenzen. innerhalb von bekannten RNS-Sequenzen. typisch 104 bis 105 Tripletts pro Protein.")
30
Mengengerüst menschliches Genom
ca Gene 1 Gen Codierung eines Proteins Größe eines Gens: ca Basenpaare, einschließlich unverstandenes „Verpackungsmaterial“ menschl. Genom ca. 2• 109 Basen = 4• 109 Gbit = 500 MB = 1CD Quelle: Mannheimer Forum 90/91, Boehringer Mannheim, darin: H.P. Vosberg: Konstanz und Variabilität im menschlichen Genom, S

33
Struktur eines Chromosoms
ca. 100 Mill. Zeichen ••• A C G G T A C ...?? ••• 1 Gen Gen Gen ••• CTG GAT CTG TAC GGG ••• ca ••• Leuzin Asparagin Leuzin ••• zugehöriges Eiweiß = Protein Aminosäuren

34
Die Buchstaben: DNS-Basen
A : Adenin C : Cytosin T : Thymin G : Guanin 1 Buchstabe = 2 bit Aminosäuren: Leuzin Asparagin • 20 Die Wörter: CTG ~ 6 bit GAT • 64 „Universeller genetischer Code“ genetische Sprache Genom (Wort der Sprache) Deutsch Faust Zucker & Insulin: Erbgut o.k. Fabrik defekt
 Deutsch Faust. Zucker & Insulin: Erbgut o.k. Fabrik defekt.")
36
Umfang des menschlichen Erbgutes
23 Chromosomen Gene (Schätzungen) 1 Chromosom Gene 1 Gen Wörter = Buchstaben = Seite Text = Kochrezept 105 Gene • 3 • 103 Buchstaben = 3 • 108 Buchstaben Umfang gesamtes Genmaterial: 1 Mill. Seiten Text = 3 Milliarden Buchstaben = 23 Bände (Chromosomen) je Seiten mit Rezepten 10 x größer ?? als informationstragendes Erbgut?
 1 Chromosom Gene. 1 Gen Wörter. = Buchstaben. = 1 Seite Text. = 1 Kochrezept. 105 Gene • 3 • 103 Buchstaben = 3 • 108 Buchstaben. Umfang gesamtes Genmaterial: 1 Mill. Seiten Text = 3 Milliarden Buchstaben. = 23 Bände (Chromosomen) je Seiten mit Rezepten. 10 x größer als informationstragendes Erbgut")
37
Variationen und Fehler
99.9 % gleich Kopierfehler Zeile fehlt, doppelt Seiten vertauscht Seite fehlt, doppelt Seite zerrissen fehlerhafte Gene haben andere Länge als richtige Längenvergleich

38
Vererbung: Chromosomen von Vater Chromosomen von Mutter falsches Rezept für Protein, z.B. für Insulin Probleme - wo? - ausschneiden - Längenvergleich Sonde = identifizierender kleiner Text-Ausschnitt ~ Positions.Ident. Enzym = genetische Schere Southern Blot = Längenvergleich

39
Genom-Projekt 3 Milliarden Buchstaben 1 $ pro Buchstabe : A C T G 3 Milliarden $ = 6% des UMTS Erlöses 2005 fertig !!! (war Schätzung 1996) massiver Einsatz von Robotern, Sommer 2000 heute schon: Hefesequenzierung Bakterienstammbaum 2020: individuelles Genoprogramm auf 1 CD
 massiver Einsatz von Robotern, Sommer heute schon: Hefesequenzierung. Bakterienstammbaum. 2020: individuelles Genoprogramm auf 1 CD.")
40
Soziale Folgen Erbkrankheiten Erkennung Vermeidung
Auswahl gesunder Embryos Wunschkinder ? Erbkrankheiten Huntington Muskeldystrophie zystische Fibrose Hypercholestrolämie Dickdarmkrebs Brustkrebs Alzheimer Multiple Sklerose Diabetes Schizophrenie Alkoholismus kriminelles Verhalten ...

41
Zukunft: Genogramm - Berufswahl Einstellung durch Arbeitgeber Versicherungen Porsche Fahrer? vorbeugende Medizin (Medikamente ?) Partnerwahl •
 Partnerwahl. •")
42
Kap. 4.4 Repräsentation von Positions-Bäumen
1. Geflecht Marke 1 Byte Zeiger 4 Bytes + Pos # 4 Bytes für Blatt a intern Expansionsfaktor 9 über Text 2. Bitvektoren + Geflecht Bis zu 64 Söhne: i interne b Blätter ... j ... repräsentiert als 128 Bits Info für vorhandene Knoten

43
00 : nicht vorhanden : Blatt : interner Knoten 11 Marke codiert durch Position in Bit Vektor für internen Knoten: Zeiger auf Bit Vektor für alle Söhne für Blatt: Positionsnummer Speicherbedarf: für Knoten mit i internen Söhnen und b Blatt-Söhnen Geflecht Bitvektoren i • 5 + b • b • 4 Bitvektor Zeiger Bitvektoren besser? i • 5+b • 5 20 i + b 4

44
Hypride Repr.: lokal entscheiden, 1 Zusatzbit pro Knoten
Weitere Idee: Pfadkontraktion a a d.h. jetzt haben alle Knoten eine Verzweigung 2 darüber Info in internen Knoten ••• b1 b1...bm ••• bm •••

45
Nochmal Bitvektoren und Geflecht
Vorschlag: Art des Knotens nicht in Bitvektor kodieren, sondern bei Knoten selbst, 1 Bit provorhendenem Knoten, z.B. Zeiger mit 31 Bits anstatt 32 Problem: Zugriff auf Info für vorhandene Knoten! Vorher war Direktzugriff auf Knoteninfo anhand des Bitvektors bestimmbar. Lösung: alle vorhandenen Söhne sequentiell abarbeiten z11, z20, z31, ... Speicherbedarf: Geflecht Bitvektoren i • 5 + b • b • 4 Vektoren besser? i • 5 + b • 9 12 + b • 4 i + b 12 5

Ähnliche Präsentationen




>")

>")
>")

>")






>")
>")

 U N I V E R S I T Ä T H A M B U R G November 2011.>")
