Präsentation herunterladen
Die Präsentation wird geladen. Bitte warten

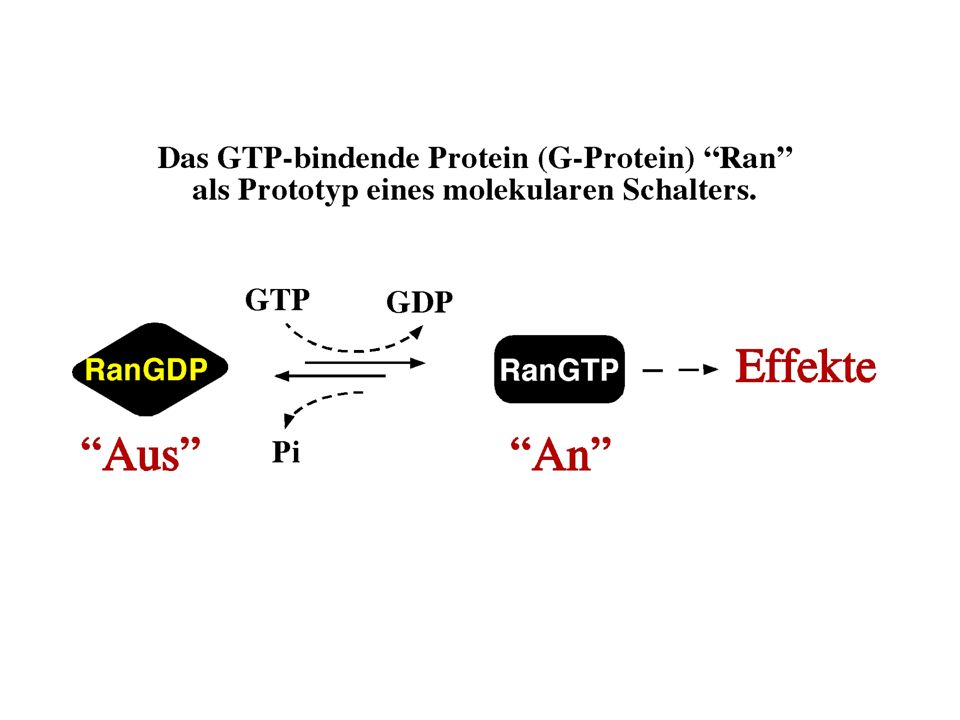
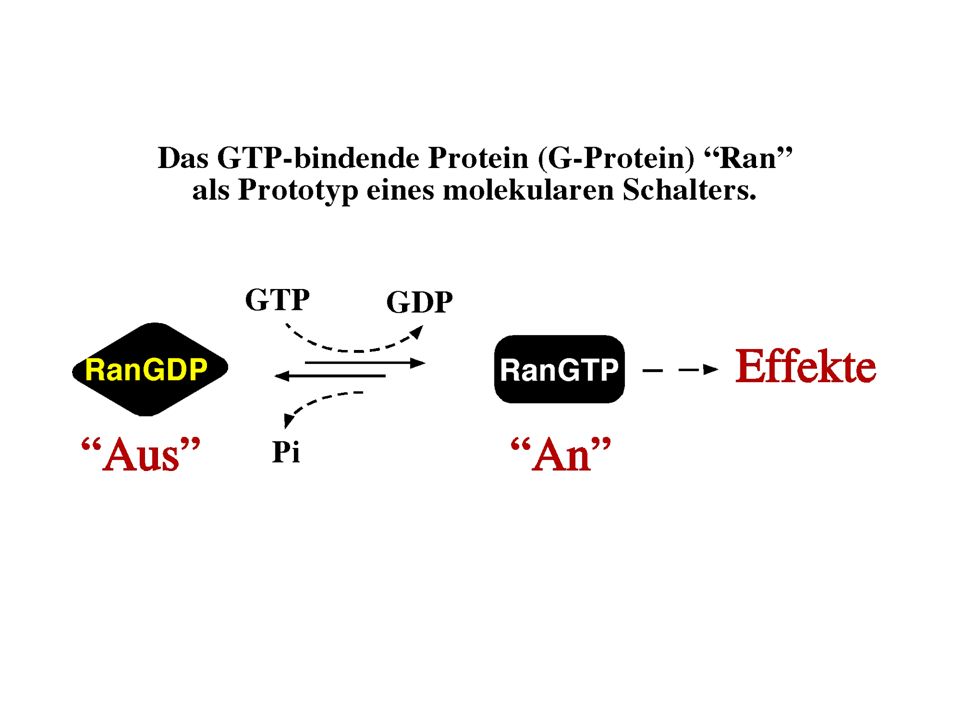
1
Zur Erinnerung... - Membranen bestehen aus unterschiedlichen Lipiden:
1. Phospholipide (Grundbaustein ist das Phosphatidat) 2. Sphingosine (Grundbaustein das Sphingosin) Von den Sphingosinen leiten sich auch die Glycolipide (Cerebroside, Ganglioside ab). 3. Cholesterin (ein Steroid) -Die Lipide sind amphipathisch. Diese Eigenschaft erlaubt den Zusammenschluss zu biologischen Doppelmembranen. -Lipid-Doppelmembranen sind für polare Moleküle nahezu impermeabel. Membranproteine vermitteln den selektiven Transport über eine Membranbarriere. -Es gibt integrale Membranproteine (Transmembrandomänen können aus a-Helices oder aus ß-Faltblättern bestehen) und periphere Membranproteine.
 2. Sphingosine (Grundbaustein das Sphingosin) Von den Sphingosinen leiten sich auch die Glycolipide (Cerebroside, Ganglioside ab). 3. Cholesterin (ein Steroid) -Die Lipide sind amphipathisch. Diese Eigenschaft erlaubt den. Zusammenschluss zu biologischen Doppelmembranen. -Lipid-Doppelmembranen sind für polare Moleküle nahezu. impermeabel. Membranproteine vermitteln den selektiven Transport. über eine Membranbarriere. -Es gibt integrale Membranproteine (Transmembrandomänen können. aus a-Helices oder aus ß-Faltblättern bestehen) und periphere. Membranproteine.")
2
Zellen höherer Lebewesen haben viele
Kompartimente ER Aufnahme über Endocytose Zellkern Golgi Sekretion Mitochondrien Lysosomen

3
Transportprozesse in der Zelle: ein generelles Problem
-Die Einteilung der Zelle in unterschiedliche Kompartimente macht biochemische Systeme notwendig, die den gerichteten Transport von Molekülen (Cargo) gewährleisten. Die wichtigsten Transportprozesse in der Zelle: -Transport Mitochondrien/Chloroplasten -Sekretion (Transport vom ER zur Zelloberfläche) -Aufnahme von Stoffen aus dem Medium (Endocytose) -Transport in kleinere Organellen wie Lysosomen (M-6-P!) -Kerntransport (nucleo-cytoplasmatischer Transport) Zellkern ist von einer Doppelmembran umschlossen!
 gewährleisten. Die wichtigsten Transportprozesse in der Zelle: -Transport Mitochondrien/Chloroplasten. -Sekretion (Transport vom ER zur Zelloberfläche) -Aufnahme von Stoffen aus dem Medium (Endocytose) -Transport in kleinere Organellen wie Lysosomen (M-6-P!) -Kerntransport (nucleo-cytoplasmatischer Transport) Zellkern ist von einer Doppelmembran umschlossen!")
4
Die Kernmembran trennt wesentliche Prozesse voneinander
Zytoplasma: Translation (benötigt tRNA, Ribosomen, mRNA) Kern: DNA Replikation, Verpackung der DNA in Chromatin, Reparatur geschädigter DNA... mRNA Synthese (Transkription, 7Methyl G-Capping, Spleißen, Poly-Adenylierung). Synthese von prä-tRNA und deren Reifung zu tRNA Synthese und Reifung von rRNA, Zusammenbau ribosomaler Untereinheiten und anderer RNA-Proteinkomplexe (RNPs).
 Kern: DNA Replikation, Verpackung der DNA in Chromatin, Reparatur geschädigter DNA... mRNA Synthese (Transkription, 7Methyl G-Capping, Spleißen, Poly-Adenylierung). Synthese von prä-tRNA und deren Reifung zu tRNA. Synthese und Reifung von rRNA, Zusammenbau ribosomaler Untereinheiten und anderer RNA-Proteinkomplexe (RNPs).")
5
Proteine „wissen“ wo sie hingehören
Ein Kernportein (z.B. ein DNA- bindendes Protein) wandert schnell nach der Synthese im Cytosol in den Kern ein cytoplasmatisches Protein (z.B. ein Enzym der Glycolyse kann nicht in den Kern)
 wandert schnell. nach der Synthese. im Cytosol in den Kern. ein cytoplasmatisches Protein. (z.B. ein Enzym der Glycolyse. kann nicht in den Kern)")
6
Was muß zwischen Kern und Zytoplasma transportiert werden?
Aktiver Import in den Kern: Sämtliche Proteine des Zellkerns (RNA- und DNA Polymerasen, Transkriptionsfaktoren, Histone, viele RNA-bindende Proteine, Spleißfaktoren), sowie ribosomale Proteine müssen importiert werden. Viele Viren, wie z.B. Influenza und HIV werden auch importiert. Aktiver Export: tRNA, ribosomale Untereinheiten, mRNA, diese werden zur Protein-biosynthese (Translation) im Zytoplasma benötigt. Die Genome vieler Viren werden zur Assemblierung der viralen Partikel aus dem Kern exportiert.
, sowie ribosomale Proteine müssen importiert werden. Viele Viren, wie z.B. Influenza und HIV werden auch importiert. Aktiver Export: tRNA, ribosomale Untereinheiten, mRNA, diese werden zur Protein-biosynthese (Translation) im Zytoplasma benötigt. Die Genome vieler Viren werden zur Assemblierung der viralen Partikel aus dem Kern exportiert.")
7
Warum ein Zellkern? Der makromolekulare Transport zwischen Kern und Zytoplasma bindet beträchtliche zelluläre Ressourcen. Warum werden die beiden Kompartimente überhaupt getrennt? Bakterien können prima ohne Zellkern leben!

8
Die Abgrenzung der genetischen Information erlaubte die Entwicklung zu höheren Organismen
Nur Eukaryonten haben in der Evolution den Sprung zu komplexen vielzelligen Organismen geschafft. Die Aufbewahrung der genetischen Information in einem spezialisierten Kompartiment trägt sicherlich zur genetischen Stabilität bei und erlaubt Eukaryonten, mehr genetische Information zu handhaben, als Prokaryonten (typischerweise das 1000-fache). Konzentrierung spezifischer Faktoren in einzelnen Kompartimenten erhöht deren lokale Konzentration und macht dadurch die katalysierten Reaktionen effektiver. Spleißen: Typische eukaryontische Gene bestehen aus Exons und Introns. Daher darf das Primärtranskript erst nach dem Spleißen translatiert werden. Die räumliche Trennung von Zellkern und Zytoplasma ist eine elegante Lösung dieses Problems. Die physische Trennung von Kern und Zytoplasma kann zur Regulation und Kontrolle der Genexpression genutzt werden.
. Konzentrierung spezifischer Faktoren in einzelnen Kompartimenten erhöht deren lokale Konzentration und macht dadurch die katalysierten Reaktionen effektiver. Spleißen: Typische eukaryontische Gene bestehen aus Exons und Introns. Daher darf das Primärtranskript erst nach dem Spleißen translatiert werden. Die räumliche Trennung von Zellkern und Zytoplasma ist eine elegante Lösung dieses Problems. Die physische Trennung von Kern und Zytoplasma kann zur Regulation und Kontrolle der Genexpression genutzt werden.")
9
Der Ort des Transports: Kernporenkomplexe (NPCs)
")
10
Doppelmembran NPC

11
From Pante &Aebi, 1997, COCB 8:397

12
Eigenschaften von Kernporenkomplexen
NPCs haben ein Masse von 125 Mda (entspricht 30 Ribosomen) und sind aus 30-50 verschiedenen Nucleoporinen aufgebaut. NPCs weisen eine 8-fache Rotationssymmetrie auf. Jedes Nukleoporin kommt daher in mindestens 8 Kopien pro Kernpore vor. Insgesamt enthält eine Kernpore etwa 1000 einzelne Polypeptide. Zahl der Kernporen pro Zellkern Hefe ≈200, menschliche Zellen ≈3000, Xenopus Oozyten ≈50 000 000.
 und sind aus verschiedenen Nucleoporinen aufgebaut. NPCs weisen eine 8-fache Rotationssymmetrie auf. Jedes Nukleoporin kommt daher in mindestens 8 Kopien pro Kernpore vor. Insgesamt enthält eine. Kernpore etwa 1000 einzelne Polypeptide. Zahl der Kernporen pro Zellkern. Hefe ≈200, menschliche Zellen ≈3000, Xenopus Oozyten ≈")
13
Transport durch die Kernpore
NPCs erlauben passive Diffusion kleiner Moleküle (praktisches Limit 20-40kDa) und aktiven Transport von Makromolekülen bis zu einem Durchmesser von 25 nm (40 nm) und einer Masse von >107 Da. Aktiver Transport benötigt metabolische Energie und wird durch spezifische Rezeptoren vermittelt. Ein und dieselbe Kernpore kann sowohl Import als auch Export vermitteln. Pro Pore und Minute werden etwa 3 ribosomale UE und 50 tRNA Moleküle exportiert und etwa 1000 Proteine importiert. Die Selektivität des aktiven Transports durch Kernporenkomplexe wird durch ein Wechselspiel von Transportsignalen und –rezeptoren erreicht. Dabei gibt es viele verschiedene Import und Exportwege durch NPCs.
 und aktiven Transport von Makromolekülen bis zu einem Durchmesser von 25 nm (40 nm) und einer Masse von >107 Da. Aktiver Transport benötigt metabolische Energie und wird durch spezifische Rezeptoren vermittelt. Ein und dieselbe Kernpore kann sowohl Import als auch Export vermitteln. Pro Pore und Minute werden etwa 3 ribosomale UE und 50 tRNA Moleküle exportiert und etwa 1000 Proteine importiert. Die Selektivität des aktiven Transports durch Kernporenkomplexe wird durch ein Wechselspiel von Transportsignalen und –rezeptoren erreicht. Dabei gibt es viele verschiedene Import und Exportwege durch NPCs.")
14
Der Kerntransport von Proteinen benötigt spezifische
Faktoren (“Importine”) ohne lösliche Transportfakoren mit Transportfaktoren
 ohne lösliche Transportfakoren. mit Transportfaktoren.")
15
Faktoren die den Import ermöglichen nennt man Importine
Ohne Importine ist der Eintritt in den Kern nicht möglich

16
Importine Importine müssen Kern- Proteine von anderen
Proteinen unterscheiden können

17
Das Kernimportsignal (nuclear localisation signal, NLS)
-Signal in biologischen Systemen muss drei Kriterien erfüllen: 1. muss decodiert werden können (Rezeptor) 2. Mutationen inaktivieren das Signal 3. Transfer der Signaleigenschaft auf ein inertes Molekül muss möglich sein Das prototypische NLS (nuclear localisation signal) stammt aus dem „large T antigen“ des Simian Virus 40: PKKKRKVE (K= Lysin, R= Arginin) eine sehr basische Sequenz!
 2. Mutationen inaktivieren das Signal. 3. Transfer der Signaleigenschaft auf ein inertes Molekül. muss möglich sein. Das prototypische NLS (nuclear localisation signal) stammt aus dem „large T antigen des Simian Virus 40: PKKKRKVE (K= Lysin, R= Arginin) eine sehr basische Sequenz!")
18
Aufreinigung der Transportfaktoren: ein biochemisches Problem

19
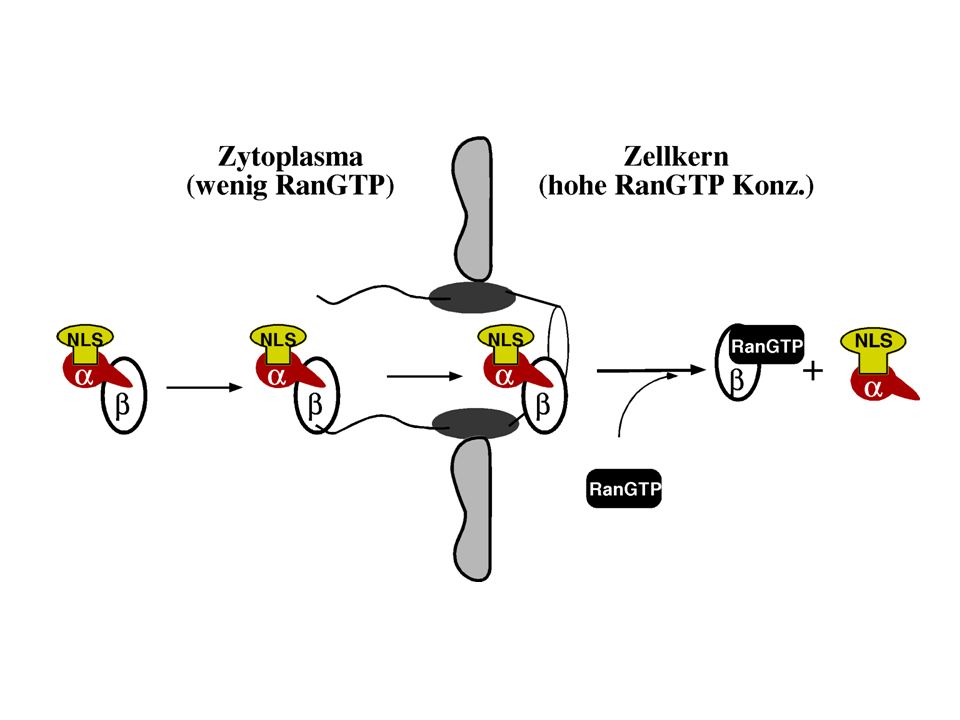
Der Rezeptor für die „klassische“ NLS Der Rezeptor ist ein Hetero-
dimer bestehend aus Importin a und Importin ß

20
Digitalis purpurea (roter Fingerhut)
")
21
Digitonin: ein Steroid aus dem roten Fingerhut
Digitonin kann spezifisch die Zellmembran zerstören, ohne den Kern zu beschädigen! herzaktive Steroide leiten sich von diesem Rest ab!

22
Wie Digitonin die Analyse des Kerntransports ermöglichte

23
Fluoreszentes NLS-Protein mit unmarkiertem Importin a/b Komplex: Importintermediate akkumulieren an den Kernporen.

24
Das Importin a/ß Dimer ist nicht für den Transport ausreichend
Docken ist möglich, nicht aber die Translokation in den Zellkern

25
Fluoreszentes NLS-protein mit Importin a/b Komplex, Ran und GTP: kompletter Import in den Kern.

26
Importin a Importin b ... beim "Docking" in Abwesenheit von Ran und GTP

27
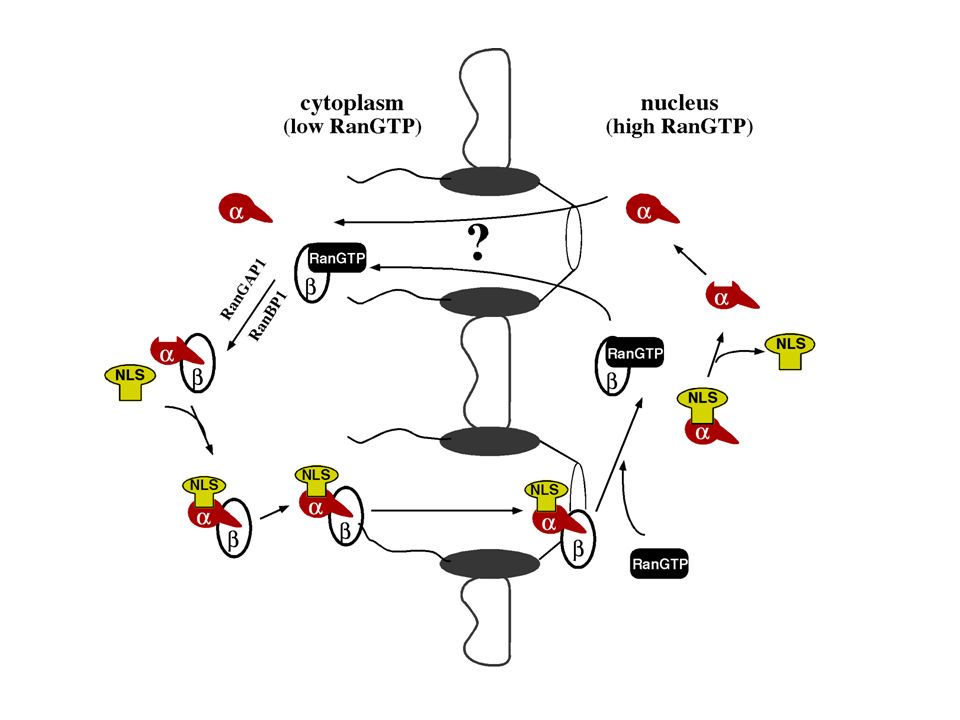
Importin a und ß trennen sich bei der Translokation
Importin b Kompletter Import in Gegenwart von Ran und GTP: die Importin Untereinheiten werden voneinander getrennt.

30
RanGTP dissoziiert den Importin a/b Komplex

31
RanGDP RanGTP RanGDP RanGTP

34
Der Kernexport von Cargo
-viele Komponenten des Zellkerns müssen auch exportiert werden (mRNAs, Ribosomen, diverse Proteine etc.) Der Export ist ebenfalls durch spezifische Signale, den Nuclear Export Signals (NES) vermittelt. LPP LER LTL die klassische NES vom HIV1 Rev Leucin-Reste sind sehr wichtig (hydrophob, vgl. NLS)
 Der Export ist ebenfalls durch spezifische Signale, den. Nuclear Export Signals (NES) vermittelt. LPP LER LTL die klassische NES vom HIV1 Rev. Leucin-Reste sind sehr wichtig. (hydrophob, vgl. NLS)")
35
Der Kernexport von Cargo
- Die NES wird im Zellkern von spezifischen Exportrezeptoren, den Exportinen erkannt. -Im Gegensatz zum Import ist die Interaktion von NES mit Exportin aber nur in Anwesenheit von RanGTP möglich. Ran GDP dissoziiert hingegen den NES-Exportin-Komplex

Ähnliche Präsentationen


>")
>")















>")